Our official English website, www.x-mol.net, welcomes your
feedback! (Note: you will need to create a separate account there.)
Phase-separation mechanism for C-terminal hyperphosphorylation of RNA polymerase II
Nature ( IF 50.5 ) Pub Date : 2018-05-30 , DOI: 10.1038/s41586-018-0174-3 Huasong Lu 1, 2 , Dan Yu 2 , Anders S Hansen 2 , Sourav Ganguly 2 , Rongdiao Liu 1 , Alec Heckert 2 , Xavier Darzacq 2 , Qiang Zhou 2
Nature ( IF 50.5 ) Pub Date : 2018-05-30 , DOI: 10.1038/s41586-018-0174-3 Huasong Lu 1, 2 , Dan Yu 2 , Anders S Hansen 2 , Sourav Ganguly 2 , Rongdiao Liu 1 , Alec Heckert 2 , Xavier Darzacq 2 , Qiang Zhou 2
Affiliation
|
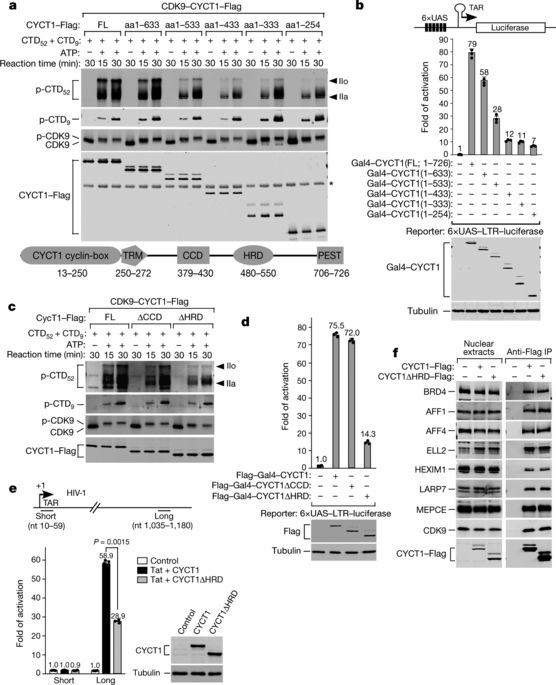
Hyperphosphorylation of the C-terminal domain (CTD) of the RPB1 subunit of human RNA polymerase (Pol) II is essential for transcriptional elongation and mRNA processing1–3. The CTD contains 52 heptapeptide repeats of the consensus sequence YSPTSPS. The highly repetitive nature and abundant possible phosphorylation sites of the CTD exert special constraints on the kinases that catalyse its hyperphosphorylation. Positive transcription elongation factor b (P-TEFb)—which consists of CDK9 and cyclin T1—is known to hyperphosphorylate the CTD and negative elongation factors to stimulate Pol II elongation1,4,5. The sequence determinant on P-TEFb that facilitates this action is currently unknown. Here we identify a histidine-rich domain in cyclin T1 that promotes the hyperphosphorylation of the CTD and stimulation of transcription by CDK9. The histidine-rich domain markedly enhances the binding of P-TEFb to the CTD and functional engagement with target genes in cells. In addition to cyclin T1, at least one other kinase—DYRK1A6—also uses a histidine-rich domain to target and hyperphosphorylate the CTD. As a low-complexity domain, the histidine-rich domain also promotes the formation of phase-separated liquid droplets in vitro, and the localization of P-TEFb to nuclear speckles that display dynamic liquid properties and are sensitive to the disruption of weak hydrophobic interactions. The CTD—which in isolation does not phase separate, despite being a low-complexity domain—is trapped within the cyclin T1 droplets, and this process is enhanced upon pre-phosphorylation by CDK7 of transcription initiation factor TFIIH1–3. By using multivalent interactions to create a phase-separated functional compartment, the histidine-rich domain in kinases targets the CTD into this environment to ensure hyperphosphorylation and efficient elongation of Pol II.The histidine-rich domain of cyclin T1 promotes phase separation into liquid droplets, which facilitates the hyperphosphorylation of the C-terminal domain repeats of RNA polymerase II.
中文翻译:

RNA聚合酶II C端过度磷酸化的相分离机制
人 RNA 聚合酶 (Pol) II RPB1 亚基的 C 末端结构域 (CTD) 的过度磷酸化对于转录延伸和 mRNA 加工至关重要 1-3。 CTD 包含共有序列 YSPTSPS 的 52 个七肽重复。 CTD 的高度重复性和丰富的可能磷酸化位点对催化其过度磷酸化的激酶施加了特殊的限制。已知正转录延伸因子 b (P-TEFb)(由 CDK9 和细胞周期蛋白 T1 组成)可使 CTD 过度磷酸化,负延伸因子可刺激 Pol II 延伸1,4,5。目前尚不清楚 P-TEFb 上促进这一作用的序列决定因素。在这里,我们在细胞周期蛋白 T1 中发现了一个富含组氨酸的结构域,它促进 CTD 的过度磷酸化并刺激 CDK9 的转录。富含组氨酸的结构域显着增强 P-TEFb 与 CTD 的结合以及与细胞中靶基因的功能结合。除了细胞周期蛋白 T1 之外,至少另一种激酶 DYRK1A6 也使用富含组氨酸的结构域来靶向 CTD 并使 CTD 过度磷酸化。作为一个低复杂性结构域,富含组氨酸的结构域还促进体外相分离液滴的形成,以及将 P-TEFb 定位到显示动态液体特性并对弱疏水相互作用的破坏敏感的核斑点上。 。 CTD(尽管是一个低复杂性结构域,但在孤立时不会发生相分离)被捕获在细胞周期蛋白 T1 液滴内,并且该过程在转录起始因子 TFIIH1-3 的 CDK7 预磷酸化后得到增强。 通过使用多价相互作用创建相分离的功能区室,激酶中的富含组氨酸结构域将 CTD 靶向到该环境中,以确保 Pol II 的过度磷酸化和有效延伸。 cyclin T1 的富含组氨酸结构域促进相分离成液滴,促进 RNA 聚合酶 II 的 C 端结构域重复的过度磷酸化。
更新日期:2018-05-30
中文翻译:
RNA聚合酶II C端过度磷酸化的相分离机制
人 RNA 聚合酶 (Pol) II RPB1 亚基的 C 末端结构域 (CTD) 的过度磷酸化对于转录延伸和 mRNA 加工至关重要 1-3。 CTD 包含共有序列 YSPTSPS 的 52 个七肽重复。 CTD 的高度重复性和丰富的可能磷酸化位点对催化其过度磷酸化的激酶施加了特殊的限制。已知正转录延伸因子 b (P-TEFb)(由 CDK9 和细胞周期蛋白 T1 组成)可使 CTD 过度磷酸化,负延伸因子可刺激 Pol II 延伸1,4,5。目前尚不清楚 P-TEFb 上促进这一作用的序列决定因素。在这里,我们在细胞周期蛋白 T1 中发现了一个富含组氨酸的结构域,它促进 CTD 的过度磷酸化并刺激 CDK9 的转录。富含组氨酸的结构域显着增强 P-TEFb 与 CTD 的结合以及与细胞中靶基因的功能结合。除了细胞周期蛋白 T1 之外,至少另一种激酶 DYRK1A6 也使用富含组氨酸的结构域来靶向 CTD 并使 CTD 过度磷酸化。作为一个低复杂性结构域,富含组氨酸的结构域还促进体外相分离液滴的形成,以及将 P-TEFb 定位到显示动态液体特性并对弱疏水相互作用的破坏敏感的核斑点上。 。 CTD(尽管是一个低复杂性结构域,但在孤立时不会发生相分离)被捕获在细胞周期蛋白 T1 液滴内,并且该过程在转录起始因子 TFIIH1-3 的 CDK7 预磷酸化后得到增强。 通过使用多价相互作用创建相分离的功能区室,激酶中的富含组氨酸结构域将 CTD 靶向到该环境中,以确保 Pol II 的过度磷酸化和有效延伸。 cyclin T1 的富含组氨酸结构域促进相分离成液滴,促进 RNA 聚合酶 II 的 C 端结构域重复的过度磷酸化。






























 京公网安备 11010802027423号
京公网安备 11010802027423号