当前位置:
X-MOL 学术
›
Nat. Commun.
›
论文详情
Our official English website, www.x-mol.net, welcomes your
feedback! (Note: you will need to create a separate account there.)
Understanding CRY2 interactions for optical control of intracellular signaling.
Nature Communications ( IF 14.7 ) Pub Date : 2017-09-15 , DOI: 10.1038/s41467-017-00648-8
Liting Duan , Jen Hope , Qunxiang Ong , Hsin-Ya Lou , Namdoo Kim , Comfrey McCarthy , Victor Acero , Michael Z. Lin , Bianxiao Cui
Nature Communications ( IF 14.7 ) Pub Date : 2017-09-15 , DOI: 10.1038/s41467-017-00648-8
Liting Duan , Jen Hope , Qunxiang Ong , Hsin-Ya Lou , Namdoo Kim , Comfrey McCarthy , Victor Acero , Michael Z. Lin , Bianxiao Cui
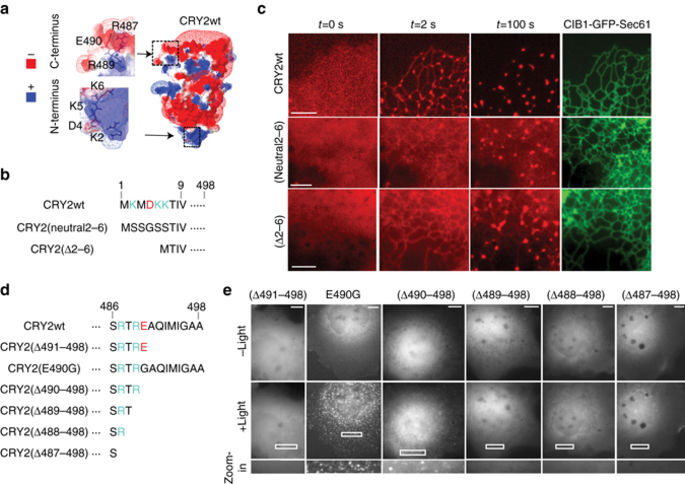
|
Arabidopsis cryptochrome 2 (CRY2) can simultaneously undergo light-dependent CRY2-CRY2 homo-oligomerization and CRY2-CIB1 hetero-dimerization, both of which have been widely used to optically control intracellular processes. Applications using CRY2-CIB1 interaction desire minimal CRY2 homo-oligomerization to avoid unintended complications, while those utilizing CRY2-CRY2 interaction prefer robust homo-oligomerization. However, selecting the type of CRY2 interaction has not been possible as the molecular mechanisms underlying CRY2 interactions are unknown. Here we report CRY2-CIB1 and CRY2-CRY2 interactions are governed by well-separated protein interfaces at the two termini of CRY2. N-terminal charges are critical for CRY2-CIB1 interaction. Moreover, two C-terminal charges impact CRY2 homo-oligomerization, with positive charges facilitating oligomerization and negative charges inhibiting it. By engineering C-terminal charges, we develop CRY2high and CRY2low with elevated or suppressed oligomerization respectively, which we use to tune the levels of Raf/MEK/ERK signaling. These results contribute to our understanding of the mechanisms underlying light-induced CRY2 interactions and enhance the controllability of CRY2-based optogenetic systems.Cryptochrome 2 (CRY2) can form light-regulated CRY2-CRY2 homo-oligomers or CRY2-CIB1 hetero-dimers, but modulating these interactions is difficult owing to the lack of interaction mechanism. Here the authors identify the interactions facilitating homo-oligomers and introduce mutations to create low and high oligomerization versions.
中文翻译:

了解CRY2相互作用对细胞内信号的光学控制。
拟南芥隐色素2(CRY2)可以同时进行光依赖性CRY2-CRY2均聚和CRY2-CIB1异二聚,这两种方法均已广泛用于光学控制细胞内过程。使用CRY2-CIB1相互作用的应用程序需要最小的CRY2均聚,以避免意外的并发症,而那些使用CRY2-CRY2相互作用的应用则更喜欢鲁棒的均聚。但是,由于CRY2相互作用的分子机制尚不清楚,因此无法选择CRY2相互作用的类型。在这里,我们报告CRY2-CIB1和CRY2-CRY2的相互作用是由在CRY2的两个末端的分离良好的蛋白质界面控制的。N端电荷对于CRY2-CIB1相互作用至关重要。此外,两个C末端电荷会影响CRY2均聚,正电荷促进低聚,负电荷抑制低聚。通过工程化C端电荷,我们分别开发了CRY2high和CRY2low寡聚体,它们的寡聚化水平较高或较低,这些均用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。我们分别开发了寡聚化水平升高或降低的CRY2high和CRY2low,可用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。我们分别开发了寡聚化水平升高或降低的CRY2high和CRY2low,可用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。
更新日期:2017-09-15
中文翻译:
了解CRY2相互作用对细胞内信号的光学控制。
拟南芥隐色素2(CRY2)可以同时进行光依赖性CRY2-CRY2均聚和CRY2-CIB1异二聚,这两种方法均已广泛用于光学控制细胞内过程。使用CRY2-CIB1相互作用的应用程序需要最小的CRY2均聚,以避免意外的并发症,而那些使用CRY2-CRY2相互作用的应用则更喜欢鲁棒的均聚。但是,由于CRY2相互作用的分子机制尚不清楚,因此无法选择CRY2相互作用的类型。在这里,我们报告CRY2-CIB1和CRY2-CRY2的相互作用是由在CRY2的两个末端的分离良好的蛋白质界面控制的。N端电荷对于CRY2-CIB1相互作用至关重要。此外,两个C末端电荷会影响CRY2均聚,正电荷促进低聚,负电荷抑制低聚。通过工程化C端电荷,我们分别开发了CRY2high和CRY2low寡聚体,它们的寡聚化水平较高或较低,这些均用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。我们分别开发了寡聚化水平升高或降低的CRY2high和CRY2low,可用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。我们分别开发了寡聚化水平升高或降低的CRY2high和CRY2low,可用于调节Raf / MEK / ERK信号传导水平。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。这些结果有助于我们理解光诱导的CRY2相互作用的机理并增强基于CRY2的光遗传系统的可控性。隐色2(CRY2)可以形成光调节的CRY2-CRY2均聚物或CRY2-CIB1异二聚体,但是由于缺乏交互机制,因此很难调节这些交互。在这里,作者确定了促进同源寡聚体的相互作用,并引入了突变以产生低聚变和高聚变形式。
































 京公网安备 11010802027423号
京公网安备 11010802027423号