Cell Death and Differentiation ( IF 13.7 ) Pub Date : 2024-10-23 , DOI: 10.1038/s41418-024-01402-6 Letizia Lanzetti
|
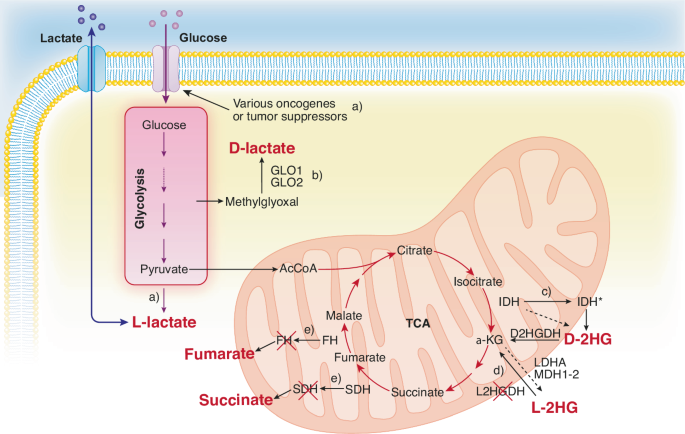
By the time a tumor reaches clinical detectability, it contains around 108–109 cells. However, during tumor formation, significant cell loss occurs due to cell death. In some estimates, it could take up to a thousand cell generations, over a ~ 20-year life-span of a tumor, to reach clinical detectability, which would correspond to a “theoretical” generation of ~1030 cells. These rough calculations indicate that cancers are under negative selection. The fact that they thrive implies that they “evolve”, and that their evolutionary trajectories are shaped by the pressure of the environment. Evolvability of a cancer is a function of its heterogeneity, which could be at the genetic, epigenetic, and ecological/microenvironmental levels [1]. These principles were summarized in a proposed classification in which Evo (evolutionary) and Eco (ecological) indexes are used to label cancers [1]. The Evo index addresses cancer cell-autonomous heterogeneity (genetic/epigenetic). The Eco index describes the ecological landscape (non-cell-autonomous) in terms of hazards to cancer survival and resources available. The reciprocal influence of Evo and Eco components is critical, as it can trigger self-sustaining loops that shape cancer evolvability [2]. Among the various hallmarks of cancer [3], metabolic alterations appear unique in that they intersect with both Evo and Eco components. This is partly because altered metabolism leads to the accumulation of oncometabolites. These oncometabolites have traditionally been viewed as mediators of non-cell-autonomous alterations in the cancer microenvironment. However, they are now increasingly recognized as inducers of genetic and epigenetic modifications. Thus, oncometabolites are uniquely positioned at the crossroads of genetic, epigenetic and ecological alterations in cancer. In this review, the mechanisms of action of oncometabolites will be summarized, together with their roles in the Evo and Eco phenotypic components of cancer evolvability. An evolutionary perspective of the impact of oncometabolites on the natural history of cancer will be presented.
中文翻译:
癌症遗传、表观遗传和生态改变十字路口的癌代谢物
当肿瘤达到临床可检测性时,它包含大约 10个 8-10 9 个细胞。然而,在肿瘤形成过程中,由于细胞死亡,会发生大量的细胞损失。在一些估计中,可能需要多达 1000 代细胞,在肿瘤的 ~ 20 年生命周期中,才能达到临床可检测性,这相当于 ~1030 个细胞的“理论”一代。这些粗略的计算表明癌症处于负选择之下。它们茁壮成长的事实意味着它们“进化”,并且它们的进化轨迹是由环境的压力塑造的。癌症的可进化性是其异质性的函数,可能在遗传、表观遗传和生态/微环境水平上 [1]。这些原则被总结在一个拟议的分类中,其中 Evo (进化) 和 Eco (生态)指数用于标记癌症 [1]。Evo 指数解决了癌细胞自主异质性(遗传/表观遗传学)的问题。生态指数描述了对癌症生存和可用资源的危害(非细胞自主)的生态景观。Evo 和 Eco 成分的相互影响至关重要,因为它可以触发塑造癌症进化性的自我维持环 [2]。在癌症的各种标志中 [3],代谢改变似乎是独一无二的,因为它们与 Evo 和 Eco 成分相交。这部分是因为新陈代谢的改变导致了 oncomtabolite 的积累。这些肿瘤代谢物传统上被视为癌症微环境中非细胞自主改变的介质。然而,它们现在越来越被认为是遗传和表观遗传修饰的诱导剂。 因此,癌代谢物在癌症遗传、表观遗传和生态改变的十字路口具有独特的位置。在本综述中,将总结肿瘤代谢物的作用机制,以及它们在癌症进化性的 Evo 和 Eco 表型成分中的作用。将介绍肿瘤代谢物对癌症自然史影响的进化观点。






























 京公网安备 11010802027423号
京公网安备 11010802027423号