Journal of Ecology ( IF 5.3 ) Pub Date : 2024-10-16 , DOI: 10.1111/1365-2745.14433 Mara Paneghel, Gil Torné, Xavier Morin, Josu G. Alday, Lluís Coll
|
1 INTRODUCTION
Mediterranean plant species exhibit a combination of life history traits that enable them to cope with fire and fire regime in different ways (Bradshaw et al., 2011; Keeley, 2012; Moris et al., 2022; Pausas, 2015). While vegetative traits contribute to the regulation of fire types and regimes—defining fire dynamics and vegetation survival—reproductive traits substantially influence auto-successional dynamics (Archibald et al., 2018; Pausas, 2015), which is paramount for the recovery of ecosystem processes after disturbances. In this context, plant species inhabiting fire-prone areas are frequently categorised based on their preferred post-disturbance reproduction strategies as either resprouters, obligate seeders or facultative seeders, which combines both (Clarke et al., 2015).
Obligate seeders rely on their seed banks and on climatic and environmental conditions following the wildfire to ensure a successful regeneration, with recruitment being a major bottleneck (Buhk et al., 2007). Often, obligate seeders possess reproductive traits that foster post-fire regeneration by enhancing either seed availability—such as through serotiny in Mediterranean conifers or fire-induced flowering—or seed germinability, which can be boosted by fire and smoke (Keeley & Pausas, 2022). Notably, serotinous species retain mature seeds in sealed cones, thus maintaining a persistent canopy seed bank until heat prompts their release (Pausas, 2015). This mechanism results in massive seed discharge following the passage of fire (Daskalakou & Thanos, 1996; Lamont et al., 1991). On the contrary, obligate seeders with soil seedbanks are generally characterised by dormant seeds, often hard coated, that rely on either fire temperatures or temperature fluctuations in the post-fire bare soil environment as a trigger for dormancy breaking (Moreira et al., 2010; Ooi et al., 2014).
Seed germination and seedling establishment after a fire are strongly contingent on the subsequent rainfall pattern (Moreno et al., 2011; Parra & Moreno, 2018). In fact, coupled with temperature, water availability defines the temporal windows during which post-fire regeneration can occur (Alvarado & Bradford, 2002; Parra & Moreno, 2018). These periods, during which seed availability matches suitable conditions for emergence, have been addressed as the window of opportunity for plant regeneration (Prévosto et al., 2014). Despite the variability in precipitation patterns (Moreno et al., 2011; Ramos, 2001) and the species-specific temperature and moisture triggers for germination (Baskin & Baskin, 2001), the temporal window of opportunity for post-fire regeneration in the Mediterranean region generally spans between autumn (September–November) and spring (March–June), and in any case when precipitations are primarily concentrated but temperatures remain mild (Prévosto et al., 2014; Quintana et al., 2004; Ramos, 2001; Santana et al., 2012). Besides, seedling emergence is most prominent in the first year after a wildfire (de Luis et al., 2008; Parra & Moreno, 2018; Quintana et al., 2004) followed by a steep decline over time (Daskalakou & Thanos, 2004; Santana et al., 2012). In particular, the most significant regeneration wave occurs during the first few months following the onset of the rainy season (Daskalakou & Thanos, 2004; Parra & Moreno, 2018). Early post-fire emergence—generally during the autumn window of opportunity—confers clear ecological advantages in terms of growth, fecundity and survival (de Luis et al., 2008). Seedlings can maximise their development throughout the rainy season (Miller et al., 2021; Santana et al., 2012; Verdú & Traveset, 2005) and enhance their chances of surviving the subsequent summer (Parra & Moreno, 2018; Vizcaíno-Palomar et al., 2014). In fact, major bottlenecks for seedling recruitments are found either shortly after emergence or due to summer drought stress—both accounting for around 50% of seedling mortality rates—hence an early emergence can be crucial for promoting establishment (Daskalakou & Thanos, 2004; Quintana et al., 2004). Despite the widespread prediction of an overall temperature increase and an escalation in intensity, duration and frequency of drought events for the Mediterranean basin (Cozannet et al., 2022; Noto et al., 2023), the potential impact of increasingly drier conditions on post-fire recruitment opportunities for Mediterranean obligate seeders remains still scarcely explored (but see Salesa et al., 2022).
Besides, climate change is likely to influence the set of climatic and weather conditions that define the wildfire niche (i.e. the combinations of weather, climatic and fuel conditions under which wildfires can occur sensu Ruffault et al., 2020), causing an extension of the wildfire season (Pausas & Keeley, 2021) and an increase in wildfire size and frequency (Dupuy et al., 2020; Ruffault et al., 2020; Turco et al., 2018; Wotton & Flannigan, 1993). For the Iberian Peninsula, the risk of fire weather is expected to increase especially in late spring and early autumn, while the historical wildfire season is concentrated in dry summer months (Calheiros et al., 2021). Fire seasonality can influence the demographic processes of post-fire regeneration at various stages, yet there is limited knowledge about the impact of changes in wildfire seasonality on post-fire tree recruitment (Miller et al., 2019; Tangney et al., 2022). For example, a shift in wildfire seasonality could alter post-fire seed dispersal patterns for serotinous species (Dunker et al., 2019), potentially creating a mismatch between seed availability and the first window of opportunity for germination. This could have cascading effects on the entire recruitment process from seed persistence in soil to the survival of emerged seedlings (Miller et al., 2019).
Whether caused by variations in the wildfire season or climate change-induced extended drought periods, the potential temporal shift or even the loss of windows of opportunity for germination and emergence could have severe consequences for post-fire recruitment. A delay in post-fire emergence, referred to as a temporal shift in emergences from the historical onset of favourable conditions, can lead to a reduction in plant fitness during the early developmental stages, particularly in terms of growth (Verdú & Traveset, 2005). In fact, smaller sizes were attributed to individuals of woody Mediterranean seeders that emerged later after a fire, although differences in size can also be related to better microsite conditions (de Luis et al., 2008) and to differences in seed mass and size, which tend to show a positive relationship with seedling height within and across species (Bladé & Vallejo, 2008; Landergott et al., 2012; Seiwa & Kikuzawa, 1991). Reduced growth could ultimately lead to lower survival because the root system could potentially not have outgrown the desiccation-sensitive soil layer by the onset of summer drought (Daskalakou & Thanos, 2004; Padilla & Pugnaire, 2007; Prévosto et al., 2014). Additionally, increased drought is likely to cause a reduction in primary productivity for recruited seedlings—those that survived the early mortality phase—especially in terms of shoot and foliar growth (Adams et al., 2015; Girard et al., 2011) and exposure to mortality hazards. Such effects could be magnified for smaller seedlings, as many ecophysiological processes are mediated by plant size (Mencuccini et al., 2005) and smaller individuals tend to be more sensitive to climatic variations (De Luis et al., 2009), especially rising temperatures (Christopoulou et al., 2022). However, thus far, the effects of a climate change-induced temporal shift in emergence time, as well as the feedback between recruit size and seedling performance and survival under drought, have been scarcely investigated in a post-fire scenario.
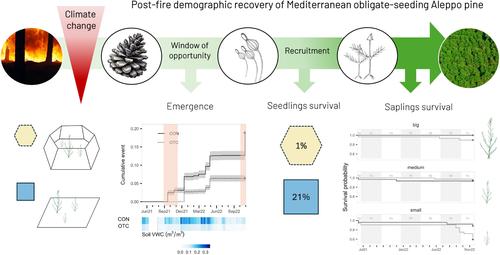
This study aimed to explore the potential impact of climate change-induced rising temperatures and seasonal variation in seed availability on post-fire regeneration dynamics of a Mediterranean obligate seeder. We selected the Aleppo pine (Pinus halepensis Mill.) as focal species, given its emblematic status as a canopy obligate seeding species in Mediterranean basin forests, producing serotinous cones that ensure massive seed release following wildfires (Daskalakou & Thanos, 1996). To achieve this, we employed an in situ experimental simulation of climate change using open-top chambers (OTCs)—a glasshouse-like setup designed primarily to increase temperature—along with a simulation of seasonal variation in seed dispersal through artificial seeding. Specifically, our objectives are to understand (i) how the increase in temperature and the seeding season influence the patterns of seedling emergence and early survival and (ii) how the increase in temperature and the size of the recruits affect the performance and survival of recruited seedlings. In relation to the first objective, we hypothesise that under a temperature increase, the autumnal window for seedling emergence will be delayed, as optimal conditions for temperature and moisture may not be met due to OTC-induced extension of summer aridity. However, we expect this autumnal delay to be followed by higher and anticipated emergences in winter, in response to milder winter temperatures under OTCs. We also expect that the temporal patterns of emergence will be more influenced by the effects of OTCs than by the seeding time, with environmental conditions such as temperature and moisture being the primary factors affecting emergence. Nevertheless, we anticipate that seed batches sown outside the historical wildfire season will show lower emergence rates due to potential mismatches between seed availability and optimal conditions for germination and emergence, posing risks to seed viability. Furthermore, we anticipate that higher temperatures within the OTCs will reduce the survival of emerged seedlings. Similarly, in relation to the second objective, we hypothesise that increased temperature will negatively affect the growth of recruited seedlings, as harsher climatic conditions can be detrimental to plant primary productivity, leading to reduced survival. We expect smaller recruits to be particularly sensitive to temperature increases, as many ecophysiological processes are mediated by plant size.
中文翻译:
温度升高威胁地中海专性播种机火灾后的自动演替动力学
1 引言
地中海植物物种表现出生活史特征的组合,使它们能够以不同的方式应对火灾和火灾状况(Bradshaw等人,2011 年;Keeley,2012 年;Moris等人,2022 年;Pausas,2015 年)。虽然营养性状有助于调节火灾类型和制度——定义火灾动态和植被生存——但生殖性状会严重影响自演替动力学(Archibald et al., 2018;Pausas,2015 年),这对于干扰后生态系统过程的恢复至关重要。在这种情况下,栖息在火灾易发地区的植物物种通常根据其首选的干扰后繁殖策略被分类为繁殖者、专性播种者或兼性播种者,两者结合在一起(Clarke等人,2015 年)。
专性播种机依靠他们的种子库以及野火后的气候和环境条件来确保成功再生,而招募是一个主要瓶颈(Buhk et al., 2007)。通常,专性播种机具有繁殖特性,可以通过提高种子的可用性(例如通过地中海针叶树中的血清化或火诱导的开花)或种子发芽性(可以通过火和烟来促进)来促进火后再生,这可以通过火和烟来增强(Keeley & Pausas,2022)。值得注意的是,血清素物种将成熟的种子保留在密封的球果中,从而维持持久的树冠种子库,直到热量促使它们释放(Pausas,2015)。这种机制导致火势过后大量种子排出(Daskalakou & Thanos,1996;Lamont et al., 1991)。相反,具有土壤种子库的专性播种机通常以休眠种子为特征,通常是硬涂层,它们依赖于火温或火后裸露土壤环境中的温度波动作为休眠打破的触发因素(Moreira等人,2010 年;Ooi et al., 2014)。
火灾后的种子发芽和幼苗建立在很大程度上取决于随后的降雨模式(Moreno et al., 2011;Parra & Moreno,2018 年)。事实上,与温度相结合,水的可用性决定了火灾后再生可以发生的时间窗口(Alvarado & Bradford,2002 年;Parra & Moreno,2018 年)。在这些时期,种子可用性与适宜的出苗条件相匹配,已被视为植物再生的机会窗口(Prévosto et al., 2014)。尽管降水模式存在变化(Moreno et al., 2011;Ramos, 2001)和物种特异性的温度和湿度触发发芽(Baskin & Baskin, 2001),地中海地区火灾后再生的机会时间窗口通常跨越秋季(9月至11月)和春季(3月至6月),以及在任何情况下降水主要集中但温度保持温和(Prévosto等人,2014;Quintana 等人,2004 年;Ramos, 2001;Santana等人,2012 年)。此外,幼苗的出苗在野火后的第一年最为突出(de Luis等人,2008 年;Parra & Moreno, 2018;Quintana等人,2004年),然后随着时间的推移急剧下降(Daskalakou & Thanos,2004年; Santana等人,2012 年)。特别是,最显著的再生浪潮发生在雨季开始后的最初几个月(Daskalakou & Thanos,2004年;Parra & Moreno,2018 年)。 火灾后的早期出现——通常是在秋季的机会之窗期间——在生长、繁殖力和生存方面赋予了明显的生态优势(de Luis et al., 2008)。幼苗可以在整个雨季最大限度地发育(Miller等人,2021 年;Santana et al., 2012;Verdú & Traveset, 2005) 并增加他们在随后的夏天存活的机会(Parra & Moreno, 2018;Vizcaíno-Palomar et al., 2014)。事实上,幼苗招募的主要瓶颈要么是在出苗后不久发现的,要么是由于夏季干旱压力——两者都占了幼苗死亡率的50%左右——因此早期出苗对于促进建立至关重要(Daskalakou & Thanos,2004;Quintana et al., 2004)。尽管人们普遍预测地中海盆地的整体温度会升高,并且干旱事件的强度、持续时间和频率会升级(Cozannet 等人,2022 年;Noto et al., 2023),但日益干燥的条件对地中海专性播种机火后招募机会的潜在影响仍很少被探索(但见 Salesa et al., 2022)。
此外,气候变化可能会影响定义野火生态位的气候和天气条件(即天气、气候和燃料条件的组合,这些条件可能会发生野火,Ruffault et al., 2020),导致野火季节的延长(Pausas & Keeley, 2021)和野火规模和频率的增加(Dupuy等人, 2020 年;Ruffault et al., 2020;Turco等人,2018 年;Wotton & Flannigan,1993 年)。对于伊比利亚半岛,预计火灾天气的风险将增加,尤其是在春末秋初,而历史上的野火季节集中在干燥的夏季(Calheiros 等人,2021 年)。火灾季节性可以在各个阶段影响火灾后再生的人口过程,但关于野火季节性变化对火灾后树木补充的影响的认识有限(Miller 等人,2019 年;Tangney et al., 2022)。例如,野火季节性的变化可能会改变血清素物种的火后种子传播模式(Dunker et al., 2019),从而可能导致种子可用性与发芽的第一个机会窗口不匹配。这可能会对从种子在土壤中的持久性到出苗存活的整个补充过程产生级联效应(Miller et al., 2019)。
无论是由野火季节的变化还是气候变化引起的长期干旱,潜在的时间变化,甚至失去发芽和出现的机会窗口,都可能对火灾后的招募产生严重后果。火灾后出苗的延迟,被称为从历史上有利条件开始的时间转变,可能导致植物在早期发育阶段的适应性降低,特别是在生长方面(Verdú & Traveset,2005)。事实上,较小的大小归因于火灾后后期出现的木质地中海播种者个体,尽管大小的差异也可能与更好的微站点条件有关(de Luis等人,2008年)以及种子质量和大小的差异,这往往显示出与物种内部和之间的幼苗高度的正相关关系(Bladé和Vallejo, 2008 年;Landergott et al., 2012;Seiwa & Kikuzawa, 1991)。生长减少最终可能导致存活率降低,因为根系在夏季干旱开始时可能无法超过对干燥敏感的土壤层(Daskalakou & Thanos,2004;Padilla & Pugnaire, 2007;Prévosto et al., 2014)。此外,干旱加剧可能会导致招募的幼苗(那些在早期死亡阶段存活下来的幼苗)的初级生产力降低,尤其是在芽和叶面生长方面(Adams等人,2015 年;Girard 等人,2011 年)和暴露于死亡风险。这种影响可以被较小的幼苗放大,因为许多生态生理过程是由植物大小介导的(Mencuccini 等人。,2005 年),而较小的个体往往对气候变化更敏感(De Luis 等人,2009 年),尤其是气温上升(Christopoulou 等人,2022 年)。然而,到目前为止,气候变化引起的出苗时间时间变化的影响,以及新兵大小和幼苗性能与干旱下存活率之间的反馈,在火灾后情景中几乎没有得到研究。
本研究旨在探讨气候变化引起的温度上升和种子供应的季节性变化对地中海专性播种机火后再生动力学的潜在影响。我们选择了阿勒颇松(Pinus halepensis Mill.)作为重点物种,因为它在地中海盆地森林中具有标志性的树冠专性种子物种的地位,可以产生血清素球果,确保在野火后大量释放种子(Daskalakou & Thanos,1996)。为了实现这一目标,我们使用开顶室 (OTC) 对气候变化进行了原位实验模拟——一种主要旨在提高温度的类似温室的设置——以及通过人工播种模拟种子传播的季节性变化。具体来说,我们的目标是了解 (i) 温度升高和播种季节如何影响幼苗出苗和早期存活模式,以及 (ii) 温度升高和新生苗的大小如何影响新生幼苗的性能和存活。关于第一个目标,我们假设在温度升高的情况下,秋季幼苗出苗的窗口将延迟,因为由于 OTC 诱导的夏季干旱延长,可能无法满足温度和湿度的最佳条件。然而,我们预计秋季延迟之后,冬季将出现更高的和预期的出苗率,以应对 OTC 下较温和的冬季温度。我们还预计,与播种时间相比,出苗的时间模式将更多地受 OTC 影响,其中温度和湿度等环境条件是影响出苗的主要因素。 尽管如此,我们预计在历史野火季节之外播种的种子批次将显示出较低的出苗率,因为种子可用性与发芽和出苗的最佳条件之间可能存在不匹配,从而对种子的生存能力构成风险。此外,我们预计 OTC 内的较高温度将降低出现的幼苗的存活率。同样,关于第二个目标,我们假设温度升高会对招募的幼苗的生长产生负面影响,因为更恶劣的气候条件可能不利于植物初级生产力,导致存活率降低。我们预计较小的新兵对温度升高特别敏感,因为许多生态生理过程是由植物大小介导的。





























 京公网安备 11010802027423号
京公网安备 11010802027423号