Journal of Ecology ( IF 5.3 ) Pub Date : 2024-08-23 , DOI: 10.1111/1365-2745.14400 Karl Andraczek 1, 2 , Laura E. Dee 3 , Alexandra Weigelt 1, 2 , Judith Hinderling 4 , Daniel Prati 4 , Gaëtane Le Provost 5, 6 , Peter Manning 5, 7 , Christian Wirth 1, 2 , Fons van der Plas 1, 8
|
1 INTRODUCTION
Global biodiversity decline (WWF, 2022) has fuelled an ongoing research debate about the consequences of biodiversity loss for ecosystem functioning (Adler et al., 2011; Dee et al., 2022; van der Plas, 2019). A focal topic in this debate is the reciprocal nature of biodiversity-productivity relationships, in which biodiversity can affect productivity but productivity also drives biodiversity (Adler et al., 2011; Grace et al., 2016). In recent decades, numerous experimental studies concluded that random biodiversity loss reduces productivity (Cardinale et al., 2012; Tilman et al., 2001). However, in non-randomly assembled (semi-)natural ecosystems (e.g. managed grasslands), these relationships are less consistent (Adler et al., 2011; Dee et al., 2023; Grace et al., 2016), which challenges the generality of biodiversity effects on productivity (van der Plas, 2019). At the same time, the direction of the causal relationship between biodiversity and productivity has been debated for decades, with many earlier studies suggesting that biodiversity can also be driven by changes in productivity (Fraser et al., 2015; Grime, 1973; Gross & Cardinale, 2007). Yet despite recent evidence suggesting that both relationships of biodiversity on productivity and of productivity on biodiversity may co-occur (Grace et al., 2016), their causal effects remain elusive (Dee et al., 2023).
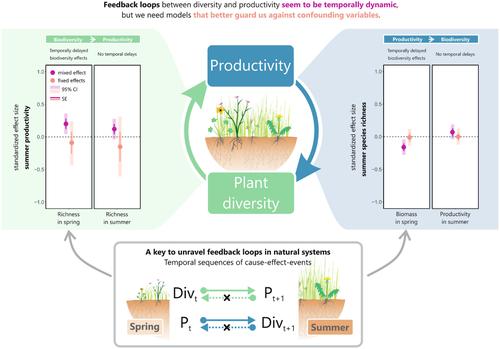
Teasing apart biodiversity-productivity relationships in natural and semi-natural ecosystems is challenging. First, confounding variables, such as land-use intensity can influence both biodiversity and productivity and modify their relationships (Grace et al., 2007; Socher et al., 2012). When unaccounted for, confounding variables can mask or mimic causal effects and lead to incorrect inferences (i.e. due to statistical bias, reviewed in Dee et al., 2023). Thus, the degree to which overall relationships in prior observational studies are positive or negative may be driven by variation in land-use intensity (Freitag et al., 2023; Grace et al., 2007; Socher et al., 2012) or how these studies control for confounding variables (Dee et al., 2023). Second, inconsistent biodiversity-productivity relationships could be caused by their reciprocal relationship (Figure 1a, Cardinale et al., 2009; Grace et al., 2007; Willig, 2011). In grasslands, high productivity can reduce plant biodiversity through increased light competition (Hautier et al., 2009) or changes in nutrient niche-dimensionality (Harpole et al., 2017). In contrast, studies focusing on effects of biodiversity on productivity often found positive relationships in artificial experimental systems (Cardinale et al., 2012; Loreau et al., 2001; Tilman et al., 2014), mainly due to increased complementarity (Barry et al., 2018), although more ‘real-world’ experiments (i.e. species loss experiments) have also found weaker relationships (Lisner et al., 2023; Sasaki et al., 2017; Smith & Knapp, 2003). These processes, however, are not mutually exclusive (Grace et al., 2016) and are hard to tease apart in observational data when they occur simultaneously (Dee et al., 2022). For instance, a negative effect of productivity on biodiversity could mask a positive effect of biodiversity on productivity (Schmid, 2002; van der Plas, 2019). Hence, to robustly estimate the effect of biodiversity on productivity and vice versa in (semi-)natural ecosystems, such as managed grasslands, isolating each direction of their reciprocal relationship from the other is crucial.

Despite the potential of reciprocal relationships between biodiversity and productivity to obscure attempts to isolate the effects of biodiversity on productivity from productivity on biodiversity and vice versa, few prior studies either attempt to account for it explicitly (but see Dee et al., 2023) or to study both directions in a single study (but see Chen et al., 2018; Grace et al., 2016; Paquette & Messier, 2011). For instance, of 31 studies reviewed in a comprehensive literature synthesis in van der Plas (2019) most do not address reverse causality (Dee et al., 2022). In a notable exception that examined effects in both directions of this bidirectional relationship, Grace et al. (2016) found a positive effect of biodiversity on productivity, and a negative effect of productivity on biodiversity in (semi-)natural grassland. However, this study used observations from single points in time, which creates challenges for determining the temporal ordering of causal effects. In particular, if productivity is inferred from plant biodiversity sampled at the same point in time (or vice versa), their effects will be hard to disentangle, causing simultaneity bias (Wooldridge, 2012)—even if they do not affect each other simultaneously. Furthermore, the effects of biodiversity on productivity (or vice versa) are likely not truly simultaneous and occur at different time scales (Costanza et al., 2007), for example, delayed effects which need a longer time to emerge (e.g. effects of biodiversity on productivity through plant–soil feedback effects). While some studies focused on long-term (across multiple years) effects between biodiversity and productivity (Isbell et al., 2018; Qiu & Cardinale, 2020), less is known about higher resolutions of temporal dynamics (e.g. within years—the scale at which productivity can reduce biodiversity through shading, for example) (Dullinger et al., 2013; Faust et al., 2012; Reich et al., 2012). However, using data from multiple time points within a year based on seasonal data allows us not only to estimate reciprocal relationships but also to explore different temporal dimensions of cause and effect (e.g. delayed causal effects from on season to another). Having repeated samples across the same sites over time (allowing analysis of within-site changes; longitudinal data hereafter) further enable statistical designs (i.e. two-way fixed-effect models; Wooldridge, 2012) that allow for control over a broader suite of observable and importantly, unobservable confounding variables without needing to know or measure them (Figure 1b,e.g. measurement error and micro-climate) (Dee et al., 2023; Larsen et al., 2019). Thus, to deal with the temporal precedence of effects and to tease out reverse causalities, longitudinal data, that is, surveys repeated both within years (across seasons) and across years, can be crucial if we aim to identify and estimate the causal effects between biodiversity and productivity (Dee et al., 2023; Larsen et al., 2019; Leszczensky & Wolbring, 2022).
Here, we compare the magnitude of the relationship of plant biodiversity on productivity with the reciprocal relationship of standing biomass (resulting from productivity) on biodiversity within managed grasslands that vary in land-use intensity. We use data collected over 2 years and two seasons per year on plant species richness, standing biomass and productivity across 150 managed grassland sites located across Germany. Previous studies observed negative correlations between biodiversity and productivity in these grasslands (Socher et al., 2012), even after controlling for multiple confounding variables (Le Provost et al., 2023). Here we build on these prior studies by using longitudinal data within a growing season, across years and so-called ‘causal inference’ methods (Figure 1b) to resolve and compare the relative magnitudes of reciprocal relationships between biodiversity and productivity. We show, for the first time, that higher-resolution longitudinal data (interannual, i.e. seasonal data, as opposed to single annual measurements used in previous studies) can be used to resolve reciprocal relationships. By using causal inference tools, we found that within managed grasslands, the estimated effects of biodiversity on productivity and vice versa are weak. When comparing inferences from our design that can control for more confounding variables to a typical statistical design in ecology (i.e. mixed-effect models), we find that our estimates differ both in signs and magnitudes, and that these effects are likely overestimated (by two standard errors) by conventional statistical designs. Ultimately, we conclude by highlighting future data needs and methods that can help us to resolve biodiversity-productivity relationships that can help to reconcile a long-running debate in ecology and ultimately, to understand how biodiversity and ecosystem functioning respond to global change.
中文翻译:
管理草原生产力与植物生物多样性之间的弱互惠关系
1 引言
全球生物多样性下降(WWF,2022 年)引发了一场关于生物多样性丧失对生态系统功能影响的持续研究辩论(Adler 等人,2011 年;Dee et al., 2022;van der Plas,2019 年)。这场辩论的一个焦点是生物多样性-生产力关系的互惠性质,其中生物多样性可以影响生产力,但生产力也会推动生物多样性(Adler 等人,2011 年;Grace et al., 2016)。近几十年来,许多实验研究得出结论,随机的生物多样性丧失会降低生产力(Cardinale et al., 2012;Tilman et al., 2001)。然而,在非随机组装的(半)自然生态系统(例如,人工管理的草原)中,这些关系不太一致(Adler et al., 2011;Dee 等人,2023 年;Grace et al., 2016),该研究挑战了生物多样性对生产力影响的普遍性(van der Plas,2019)。与此同时,生物多样性与生产力之间因果关系的方向已经争论了几十年,许多早期研究表明,生物多样性也可以由生产力的变化驱动(Fraser et al., 2015;Grime,1973 年;Gross & Cardinale,2007 年)。然而,尽管最近的证据表明,生物多样性与生产力的关系和生产力与生物多样性的关系可能同时发生(Grace et al., 2016),但它们的因果效应仍然难以捉摸(Dee et al., 2023)。
梳理自然和半自然生态系统中的生物多样性与生产力的关系是具有挑战性的。首先,混杂变量,如土地利用强度,可以影响生物多样性和生产力,并改变它们之间的关系(Grace et al., 2007;Socher et al., 2012)。如果不加以解释,混杂变量可能会掩盖或模仿因果效应并导致不正确的推断(即由于统计偏差,Dee 等人,2023 年综述)。因此,先前观察研究中总体关系是正或负的程度可能由土地利用强度的变化驱动(Freitag et al., 2023;Grace et al., 2007;Socher et al., 2012) 或这些研究如何控制混杂变量(Dee et al., 2023)。其次,生物多样性-生产力关系不一致可能是由它们的互惠关系引起的(图 1a,Cardinale 等人,2009 年;Grace et al., 2007;Willig,2011 年)。在草原中,高生产力可以通过增加光竞争(Hautier et al., 2009)或改变养分生态位维度(Harpole et al., 2017)来减少植物生物多样性。相比之下,关注生物多样性对生产力影响的研究经常发现人工实验系统中的正相关关系(Cardinale et al., 2012;Loreau等人,2001 年;Tilman等人,2014 年),主要是由于互补性增加(Barry等人,2018 年),尽管更多的“真实世界”实验(即物种丧失实验)也发现了较弱的关系(Lisner等人,2023 年;Sasaki等人,2017 年;Smith & Knapp,2003 年)。 然而,这些过程并不是相互排斥的(Grace et al., 2016),并且当它们同时发生时,很难在观测数据中区分(Dee et al., 2022)。例如,生产力对生物多样性的负面影响可能会掩盖生物多样性对生产力的积极影响(Schmid,2002;van der Plas,2019)。因此,要稳健地估计生物多样性对生产力的影响,反之亦然,在(半)自然生态系统(例如人工草原)中,将它们互惠关系的每个方向与另一个方向隔离开来是至关重要的。
在图窗查看器PowerPoint 中打开
(a) 显示植物生物多样性 (B) 和生产力 (P) 之间的互惠关系的概念模型,以及为(半)自然系统假设的其他可观察的混杂因素(即环境和干扰)。(b) 利用生物多样性和生产力的时间序列进行统计设计,以解开互惠关系,并控制可观察和不可观察的混杂因素(如土壤肥力、气候和土地利用)。(c) (半)自然系统(如农业管理的草原)中生物多样性和生产力的预期变化。箭头的大小表示先前来自(农业管理的)草原的发现所暗示的关系的预期相对重要性(Grace et al., 2007;Socher et al., 2012)。请注意,所描述的生物多样性和生产力的假设变化是基于经典生物多样性实验的结果,并且在更“真实世界”的设置下可能会有所不同(见第 4 节)。
尽管生物多样性和生产力之间的互惠关系可能会掩盖将生物多样性对生产力的影响与生产力对生物多样性的影响区分开来的尝试,反之亦然,但很少有以前的研究试图明确解释它(但见 Dee 等人,2023 年)或在单一研究中研究两个方向(但见 Chen 等人, 2018 年;Grace et al., 2016;Paquette & Messier, 2011)。例如,在 van der Plas (2019) 的综合文献综合中回顾的 31 项研究中,大多数没有涉及反向因果关系(Dee et al., 2022)。Grace 等人(2016 年)发现,在(半)自然草原中,生物多样性对生产力有积极影响,而生产力对生物多样性有消极影响。然而,这项研究使用了来自单个时间点的观察结果,这为确定因果效应的时间顺序带来了挑战。特别是,如果从同一时间点采样的植物生物多样性中推断出生产力(反之亦然),它们的影响将很难理清,从而导致同时性偏差(Wooldridge,2012)——即使它们不会同时相互影响。此外,生物多样性对生产力的影响(反之亦然)可能不是真正同时发生的,而是在不同的时间尺度上发生的(Costanza et al., 2007),例如,需要较长时间才能显现的延迟效应(例如,生物多样性通过植物-土壤反馈效应对生产力的影响)。虽然一些研究侧重于生物多样性和生产力之间的长期(跨越多年)影响(Isbell 等人。,2018 年;Qiu & Cardinale,2020),但对于时间动态的更高分辨率(例如,几年内——生产力可以通过阴影减少生物多样性的规模)(Dullinger等人,2013年;Faust et al., 2012;Reich et al., 2012)。然而,根据季节数据使用一年内多个时间点的数据,不仅可以让我们估计互惠关系,还可以探索因果关系的不同时间维度(例如,从季节到另一个季节的延迟因果效应)。随着时间的推移,在同一地点重复采样(允许分析地点内的变化;下文为纵向数据)进一步使统计设计成为可能(即双向固定效应模型;Wooldridge,2012 年),允许控制更广泛的可观察且重要的是,不可观察的混杂变量,而无需了解或测量它们(图 1b,例如测量误差和微气候)(Dee 等人,2023 年;Larsen等人,2019 年)。因此,为了处理影响的时间优先性并梳理出反向因果关系,如果我们的目标是识别和估计生物多样性与生产力之间的因果效应,那么纵向数据,即在几年内(跨季节)和跨几年重复的调查可能至关重要(Dee 等人,2023 年;Larsen等人,2019 年;Leszczensky & Wolbring,2022 年)。
在这里,我们比较了植物生物多样性与生产力的关系大小,以及土地利用强度不同的管理草原内站立生物量(由生产力产生)对生物多样性的互惠关系。我们使用 2 年和每年 2 个季节收集的数据,这些数据涉及德国 150 个管理草原的植物物种丰富度、站立生物量和生产力。以前的研究观察到这些草原的生物多样性与生产力之间存在负相关(Socher et al., 2012),即使在控制了多个混杂变量之后(Le Provost et al., 2023)。在这里,我们以这些先前的研究为基础,使用生长季节、跨年的纵向数据和所谓的“因果推理”方法(图 1b)来解决和比较生物多样性和生产力之间互惠关系的相对大小。我们首次表明,更高分辨率的纵向数据(年际数据,即季节性数据,而不是以前研究中使用的单个年度测量)可用于解决互惠关系。通过使用因果推理工具,我们发现在受管理的草原中,生物多样性对生产力的估计影响很弱,反之亦然。当将我们的设计中可以控制更多混杂变量的推论与生态学中的典型统计设计(即混合效应模型)进行比较时,我们发现我们的估计值在符号和幅度上都不同,并且这些效应可能被传统统计设计高估(两个标准误差)。 最后,我们强调了可以帮助我们解决生物多样性与生产力关系的未来数据需求和方法,这有助于调和生态学中长期存在的争论,并最终了解生物多样性和生态系统功能如何应对全球变化。






























 京公网安备 11010802027423号
京公网安备 11010802027423号