Leukemia ( IF 12.8 ) Pub Date : 2024-07-30 , DOI: 10.1038/s41375-024-02349-w Xueshuai Han 1, 2, 3 , Jingru Sui 1, 2, 3 , Kui Nie 4 , Yang Zhao 1, 2 , Xuan Lv 1, 2, 3 , Jindou Xie 1, 2, 3 , Leonard Tan 5 , Rex K H Au-Yeung 6 , Jiao Ma 7 , Giorgio Inghirami 4 , Olivier Elemento 8 , Wayne Tam 4, 9 , Zhaoqi Liu 1, 2, 3
|
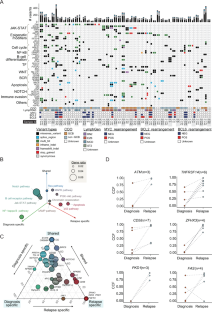
Diffuse large B-cell lymphoma (DLBCL) is the most common form of non-Hodgkin lymphoma, and over 60% patients can achieve durable remission after receiving standard immunotherapy treatment that comprises rituximab, cyclophosphamide, doxorubicin, vincristine, and prednisolone (R-CHOP) [1,2,3]. The remaining patients are refractory to this frontline treatment or suffer tumor relapses with an extremely poor prognosis [4]. Due to the limited understanding of the molecular pathogenesis of DLBCL relapse, there are currently no effective approaches to overcome resistance of R-CHOP. Therefore, characterization of the clonal evolutionary pattern and relapse-specific lesions accounting for drug resistance in relapsed DLBCL are vital for finding new treatment solutions. However, due to the difficulty in obtaining relapse samples, previous studies to understand the genetic changes upon disease recurrence are usually limited by the small number of cases with paired diagnosis-relapse sampling [5]. To characterize the evolutionary landscape of DLBCL under treatment, in this study, we performed whole-exome sequencing (WES) of matched diagnosis and relapse DNA samples from 44 DLBCL patients receiving R-CHOP treatment, together with deep targeted sequencing of 481 candidate genes in 26 patients (Table S1, Supplementary Fig. 1A).
Genomic variants emerging in tumor relapse or expanding under therapy may account for treatment failure. Hence, we used three complementary approaches to identify relapse-associated variants. Firstly, we compared the mutation frequency between the diagnosis-relapse pairs in our cohort. We found that most of the known DLBCL drivers have similar mutation frequencies (Fig. 1C). Notably, we identified several genes appeared to be mutated exclusively in recurrent tumors, including ZFHX3 (4/44) and PKD1 (3/44). ZFHX3, which encodes a transcription factor that negatively regulates c-Myb and transactivates CDKN1A, has been reported as a tumor suppressor gene in several cancers [7]. PKD1 is a serine/threonine kinase involved in a multitude of mechanisms associated with tumor progression, such as invasion, proliferation, and apoptosis [8]. Secondly, we compared the mutation frequency from our relapse DLBCL with three independent primary DLBCL cohorts (in total 1861 cases) [6, 9, 10]. By this comparison, 129 genes show a significantly higher mutation frequency in our relapsed tumors, including CD58 and ZFHX3 (Supplementary Fig. 5A, Table S4). CD58 activates T cells and natural killer (NK) cells by binding to CD2 receptors. The absence of CD58 may allow DLBCL cells to escape NK and T cell-mediated immune surveillance [11]. Finally, to characterize the evolutionary pattern of mutations under treatment, we compared the mutation Cancer Cell Fraction (CCF) between DLBCL primary and relapsed samples. A higher CCF at relapse was observed for a number of mutations involved in immune escape (CD58, TNFRSF14, FAS), apoptosis (ATM, PKD1) or transcriptional regulation (ZFHX3), suggesting new acquirements of mutations or clonal expansion at tumor relapse (Fig. 1D and Supplementary Fig. 5B). Loss of ATM was reported to lead to poor treatment response and resistance to DNA damaging agents in B-cell malignancies [12]. Interestingly, we found that TNFRSF14 alterations were specifically enriched in relapse GCB-DLBCL (n = 6) (Supplementary Fig. 5C). Consistently, in a public primary DLBCL cohort, TNFRSF14 alterations were more common in GCB-DLBCL than in ABC-DLBCL or unclassified DLBCL (Supplementary Fig. 5D) [6]. Notably, 80% of TNFRSF14 mutations were predicted to be loss-of-function mutations (Table S5). In addition, mutations in TNFRSF14 (p = 0.039) were associated with shorter overall survival in GCB-DLBCL (Supplementary Fig. 5E) [6]. These results support the hypothesis that TNFRSF14 alterations define a malignant subtype of GCB-DLBCL.
中文翻译:
肿瘤进化分析揭示了弥漫性大 B 细胞淋巴瘤复发中的免疫逃逸相关突变
弥漫性大 B 细胞淋巴瘤 (DLBCL) 是最常见的非霍奇金淋巴瘤,超过 60% 的患者在接受标准免疫疗法(包括利妥昔单抗、环磷酰胺、阿霉素、长春新碱和泼尼松龙 (R-CHOP) 治疗后)可实现持久缓解)[1,2,3]。其余患者对该一线治疗耐药或肿瘤复发,预后极差[4]。由于对DLBCL复发的分子发病机制了解有限,目前尚无有效的方法来克服R-CHOP的耐药性。因此,克隆进化模式的表征和复发性 DLBCL 耐药性的复发特异性病变对于寻找新的治疗解决方案至关重要。然而,由于获得复发样本的困难,先前了解疾病复发时基因变化的研究通常受到配对诊断-复发采样病例数量较少的限制[5]。为了描绘治疗中 DLBCL 的进化景观,在本研究中,我们对 44 名接受 R-CHOP 治疗的 DLBCL 患者的匹配诊断和复发 DNA 样本进行了全外显子组测序 (WES),并对 481 个候选基因进行了深度靶向测序。 26 名患者(表 S1,补充图 1A)。
肿瘤复发或治疗期间扩大时出现的基因变异可能是治疗失败的原因。因此,我们使用了三种互补的方法来识别与复发相关的变异。首先,我们比较了队列中诊断-复发对之间的突变频率。我们发现大多数已知的 DLBCL 驱动因素具有相似的突变频率(图 1C)。值得注意的是,我们发现了几个基因似乎只在复发性肿瘤中发生突变,包括ZFHX3 (4/44) 和PKD1 (3/44)。 ZFHX3编码负向调节 c-Myb 并反式激活CDKN1A的转录因子,已被报道为多种癌症中的肿瘤抑制基因 [7]。 PKD1是一种丝氨酸/苏氨酸激酶,参与与肿瘤进展相关的多种机制,例如侵袭、增殖和凋亡 [8]。其次,我们将复发性 DLBCL 与三个独立的原发性 DLBCL 队列(总共 1861 例)的突变频率进行了比较 [6,9,10]。通过比较,129 个基因在我们的复发肿瘤中显示出显着更高的突变频率,包括CD58和ZFHX3 (补充图 5A,表 S4)。 CD58 通过与 CD2 受体结合来激活 T 细胞和自然杀伤 (NK) 细胞。 CD58 的缺失可能会让 DLBCL 细胞逃避 NK 和 T 细胞介导的免疫监视 [11]。最后,为了表征治疗下突变的进化模式,我们比较了 DLBCL 原发样本和复发样本之间的突变癌细胞分数 (CCF)。 观察到许多涉及免疫逃逸( CD58 、 TNFRSF14 、 FAS )、细胞凋亡( ATM、PKD1 )或转录调节( ZFHX3 )的突变在复发时具有较高的CCF,这表明肿瘤复发时出现了新的突变或克隆扩张(图) .1D 和补充图 5B)。据报道,ATM 的缺失会导致 B 细胞恶性肿瘤的治疗反应不佳和对 DNA 损伤剂产生耐药性[12]。有趣的是,我们发现TNFRSF14 的改变在复发性 GCB-DLBCL 中特别丰富( n = 6)(补充图 5C)。一致的是,在公共原发性 DLBCL 队列中,GCB-DLBCL 中的TNFRSF14改变比 ABC-DLBCL 或未分类的 DLBCL 中更常见(补充图 5D)[6]。值得注意的是,80% 的TNFRSF14突变被预测为功能丧失突变(表 S5)。此外, TNFRSF14突变 ( p = 0.039) 与 GCB-DLBCL 较短的总生存期相关(补充图 5E)[6]。这些结果支持以下假设: TNFRSF14改变定义了 GCB-DLBCL 的恶性亚型。






























 京公网安备 11010802027423号
京公网安备 11010802027423号