Journal of Ecology ( IF 5.3 ) Pub Date : 2024-07-29 , DOI: 10.1111/1365-2745.14380 Julie A. Tierney 1 , Nina Wurzburger 2
|
1 INTRODUCTION
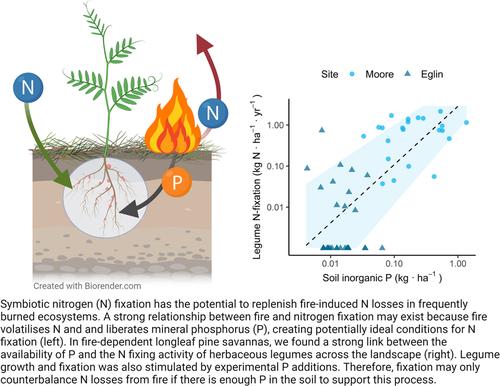
The frequent fires that maintain savanna landscapes induce large, repeated losses of nitrogen (N), a soil nutrient that commonly limits plant growth (Carter & Foster, 2004; Tierney et al., 2019; Wan et al., 2001). Consequently, fire-dependent ecosystems are thought to be N-poor. Plants that symbiotically associate with N-fixing bacteria (hereafter, N fixers) can provide a natural mechanism to replenish fire-driven N losses. As a result, a strong relationship between fire and fixation is thought to exist because fire volatilizes N, creating N-poor soils that favour plants with the N-fixing strategy. The link between fire and fixation may be further reinforced by the fact that fire mineralizes phosphorus (P) (Boring et al., 2004), a soil nutrient that commonly limits N fixers and fixation (Ament et al., 2018; Tognetti et al., 2021; Vitousek & Howarth, 1991). However, if a fire-dependent ecosystem is P-poor—such as those that occur on old, highly weathered soils (Walker & Syers, 1976)—it is unclear whether the amount of P in soil and vegetation that is available for thermal mineralization is sufficient to support fixation. If P limitation prevents N fixers from replacing N losses from fire, this can cause the decline of ecosystem N stocks over time (Pellegrini et al., 2017), potentially reducing the ecosystem's ability to recover biomass after subsequent fire events.
A case study of this phenomenon may be found in the sandhill longleaf pine savannas of the south-eastern United States. These fire-dependent ecosystems house a diversity and abundance of herbaceous legumes (Fabaceae) capable of symbiotically fixing N (Hainds et al., 1999; Norden & Kirkman, 2006). Although fire stimulates the growth of herbaceous legumes native to longleaf pine savannas (Hiers & Mitchell, 2007), a large-scale field study indicated that these legumes rarely possess N-fixing root nodules, and only fix enough N to replace 0.25%–3% of the N required to balance annual ecosystem losses from fire (Tierney et al., 2019).
While longleaf pine savannas have historically been categorized as extremely N-poor (Wilson et al., 1999, 2002), recent human influences have likely enriched these systems with N, potentially altering the relationship between fire, P and N fixation. Anthropogenic N deposition and fertilizer application in agriculture have dramatically increased the amount of reactive N in terrestrial ecosystems (Galloway et al., 2008; Gerber et al., 2013; Lamarque et al., 2013). This N enrichment has the potential to reduce the ecological importance of N fixation to replace N losses from fire, and could shift N limitation of primary productivity towards P limitation (Vitousek et al., 2010). In support of the idea that N availability is sufficient in longleaf pine savannas, long-term additions of fertilizer N did not stimulate productivity (Kirkman et al., 2016). Furthermore, another study found that rates of N mineralization appeared more than sufficient to support both tree and understorey plant growth, even in young stands undergoing rapid biomass growth (Tierney et al., 2019).
If contemporary longleaf pine ecosystems have sufficient available N, the observation that N fixation fails to replenish fire-driven N losses (Tierney et al., 2019) may be explained by two non-mutually exclusive mechanisms—N sufficiency suppresses fixation, or P limitation constrains fixation. Although losses of P via leaching and post-fire ash transport are substantially lower than those of N (Bodí et al., 2014; Hedin et al., 2003; Johnson et al., 2007; Lavoie et al., 2010), N can be replenished via N fixation while P is a finite soil resource derived from weathered rock. Ecosystem stocks of P are largely dictated by parent material and soil age (Walker & Syers, 1976), and the soils underlying the sandhill longleaf pine savannas are coarse, Cretaceous-age sediments that are extremely depauperate in P (Peet, 2006). While rare, field fertilization studies have indicated that P availability can limit longleaf pine growth (Dickens, 2001; Lewis, 1977), and one study observed that P addition increased the biomass of herbaceous legumes (Brewer & Cralle, 2003). Limitation by P may be further enhanced in the modern longleaf pine landscape if N is indeed in excess.
Legumes fix N with rhizobia, but they can also simultaneously form symbiotic relationships with arbuscular mycorrhizal fungi (AMF) to acquire N or P. Many studies have found a synergistic effect between these two below-ground symbionts, where AMF colonization stimulates legume growth, enhances P uptake from soil and increases N fixation activity (Chalk et al., 2006; Hack et al., 2019). In cases where P limits N fixation, legumes may invest more in AMF to acquire P. Moreover, legumes may also harness fixed N to construct extracellular phosphatases that liberate P in soil and enhance P availability (Houlton et al., 2008). Understanding the dynamics between these below-ground symbiotic relationships and soil nutrient availability is critical for elucidating the factors that constrain the ability of N fixers to rehabilitate the N cycle after fire.
Here, we evaluated how soil N and P may modulate the relationship between fire and N fixation in sandhill longleaf pine savannas, where we previously documented that N fixation fails to replenish fire-driven N losses. We were motivated by the question: does N sufficiency suppress, or P availability constrain, the ability of legumes to balance N losses from fire? We employed a combined observational and experimental approach to address this question. First, we conducted an observational study to investigate if and how soil N and P availability influences legume fixation across 54 plots of sandhill longleaf pine across gradients of fire return interval (1.5–20 years) and stand age (2.5 to at least 227 years). We also further explored the relationship between nutrient availability and biological nutrient acquisition by quantifying extracellular enzymes used to liberate N and P from organic matter, a reflection of the microbial demand for these nutrients. Subsequently, we conducted a nutrient manipulation experiment to more directly investigate the effects of N and P addition on legume growth, N fixation, foliar chemistry and AMF colonization in longleaf pine savannas.
中文翻译:
磷控制依赖火的长叶松稀树草原中的共生固氮
1 简介
维持稀树草原景观的频繁火灾会导致氮 (N) 的大量、反复损失,氮是一种通常限制植物生长的土壤养分(Carter & Foster, 2004 ;Tierney 等, 2019 ;Wan 等, 2001 )。因此,依赖火的生态系统被认为是贫氮的。与固氮细菌(以下简称固氮菌)共生的植物可以提供一种自然机制来补充火灾造成的氮损失。因此,人们认为火和固氮之间存在着密切的关系,因为火会挥发氮,形成贫氮土壤,有利于植物进行固氮策略。火使磷 (P) 矿化(Boring 等人, 2004 年),磷是一种通常限制氮固定剂和固定作用的土壤养分(Ament 等人, 2018 年;Tognetti 等人),这一事实可能会进一步加强火与固定之间的联系(Ament 等人,2018 年) ., 2021 ;维图塞克和豪沃斯, 1991 )。然而,如果一个依赖火的生态系统是贫磷的——比如那些发生在古老的、高度风化的土壤上的生态系统(Walker & Syers, 1976 )——那么就不清楚土壤和植被中可用于热矿化的磷含量是否足够。足以支持固定。如果磷限制阻止固氮剂补充火灾造成的氮损失,这可能会导致生态系统氮储量随着时间的推移而下降(Pellegrini 等, 2017 ),可能会降低生态系统在后续火灾事件后恢复生物量的能力。
这种现象的案例研究可以在美国东南部的沙丘长叶松稀树草原中找到。这些依赖火的生态系统拥有多样性和丰富的草本豆科植物(豆科),能够共生固定氮(Hainds 等, 1999 ;Norden 和 Kirkman, 2006 )。虽然火灾刺激了长叶松稀树草原原生的草本豆科植物的生长(Hiers & Mitchell, 2007 ),但大规模的实地研究表明这些豆科植物很少具有固氮根瘤,并且仅固定足够的氮来替代 0.25%–3平衡火灾导致的年度生态系统损失所需的氮百分比(Tierney 等人, 2019 )。
虽然长叶松稀树草原历来被归类为氮元素极度匮乏的地区(Wilson 等人, 1999 年, 2002 年),但最近人类的影响可能使这些系统富含氮元素,从而可能改变火、磷和氮固定之间的关系。农业中的人为氮沉降和施肥显着增加了陆地生态系统中活性氮的数量(Galloway等, 2008 ;Gerber等, 2013 ;Lamarque等, 2013 )。这种氮富集有可能降低固氮的生态重要性,以取代火灾造成的氮损失,并可能将初级生产力的氮限制转变为磷限制(Vitousek等, 2010 )。为了支持长叶松稀树草原中氮肥供应充足的观点,长期添加氮肥并没有刺激生产力(Kirkman 等, 2016 )。此外,另一项研究发现,氮矿化率似乎足以支持树木和林下植物的生长,即使是在生物量快速增长的幼林中(Tierney et al., 2019 )。
如果当代长叶松生态系统有足够的可用氮,则氮固定无法补充火灾导致的氮损失的观察结果(Tierney等人, 2019 )可以用两种非互斥机制来解释:氮充足抑制固定,或磷限制限制固定。尽管磷通过淋滤和火后灰烬运输造成的损失远低于氮(Bodí et al., 2014 ;Hedin et al., 2003 ;Johnson et al., 2007 ;Lavoie et al., 2010 ),N可以通过固氮来补充,而磷是来自风化岩石的有限土壤资源。生态系统的磷储量很大程度上取决于母质和土壤年龄(Walker & Syers, 1976 ),沙丘长叶松稀树草原下的土壤是粗糙的白垩纪沉积物,磷含量极其贫乏(Peet, 2006 )。虽然很少见,但田间施肥研究表明磷的有效性会限制长叶松的生长(Dickens, 2001 ;Lewis, 1977 ),一项研究观察到磷的添加增加了草本豆科植物的生物量(Brewer&Cralle, 2003 )。如果氮确实过量,在现代长叶松景观中,磷的限制可能会进一步增强。
豆科植物通过根瘤菌固定氮,但它们也可以同时与丛枝菌根真菌 (AMF) 形成共生关系以获得氮或磷。许多研究发现这两种地下共生体之间存在协同效应,其中 AMF 定殖刺激豆科植物生长,增强豆科植物生长。从土壤中吸收磷并增加固氮活性(Chalk等, 2006 ;Hack等, 2019 )。在磷限制氮固定的情况下,豆类可能会在 AMF 上投入更多资金来获取磷。此外,豆类还可以利用固定的氮来构建细胞外磷酸酶,从而释放土壤中的磷并提高磷的利用率(Houlton 等, 2008 )。了解这些地下共生关系与土壤养分可用性之间的动态对于阐明限制固氮剂在火灾后恢复氮循环能力的因素至关重要。
在这里,我们评估了土壤氮和磷如何调节沙丘长叶松稀树草原中火灾和氮固定之间的关系,我们之前记录了氮固定无法补充火灾导致的氮损失。我们的动机是这样一个问题:氮的充足性是否会抑制或磷的可用性限制豆科植物平衡火灾造成的氮损失的能力?我们采用观察和实验相结合的方法来解决这个问题。首先,我们进行了一项观察性研究,以调查土壤氮和磷的有效性是否以及如何影响 54 个沙丘长叶松地块的豆科植物固定,这些地块在火返间隔(1.5-20 年)和林龄(2.5 至至少 227 年)的梯度范围内。 。我们还通过量化用于从有机物中释放氮和磷的细胞外酶(反映了微生物对这些营养物的需求),进一步探讨了营养物可用性和生物营养物获取之间的关系。随后,我们进行了营养操纵实验,以更直接地研究氮和磷的添加对长叶松稀树草原中豆科植物生长、固氮、叶化学和 AMF 定植的影响。






























 京公网安备 11010802027423号
京公网安备 11010802027423号