Journal of Ecology ( IF 5.3 ) Pub Date : 2024-07-12 , DOI: 10.1111/1365-2745.14360 Lena Sachsenmaier 1, 2 , Florian Schnabel 1, 2, 3 , Peter Dietrich 4 , Nico Eisenhauer 1, 5 , Olga Ferlian 1, 5 , Julius Quosh 1, 5 , Ronny Richter 1, 2 , Christian Wirth 1, 2, 6
|
1 INTRODUCTION
Global warming leads to an increased likelihood of severe and consecutive droughts (Hari et al., 2020; Spinoni et al., 2018). Forest ecosystems, in particular, face threats from these droughts, given the long generation time and slow growth of trees compared to other plants (Brodribb et al., 2020). This is of great concern because, in addition to preserving biodiversity conservation and other ecosystem services, forests play a crucial role as carbon sinks, thus contributing to the mitigation of present and future climate change (Anderegg et al., 2020).
In 2018, Northern and Central Europe experienced an extraordinary compound drought, characterized by insufficient precipitation and heatwaves (Zscheischler et al., 2020; Zscheischler & Fischer, 2020). The exceptionally dry soil conditions persisted in 2019 and in vast areas of Central Europe, they lasted even through the year of 2020 (Rakovec et al., 2022). The occurrence of these three consecutive drought years from 2018 to 2020 (hereafter referred to as the ‘2018–2020 drought’) marks an unprecedented drought situation in Central Europe, at least within the last 250 years (Bastos et al., 2021; Hari et al., 2020; Rakovec et al., 2022). Such compound and consecutive droughts are substantially increasing in frequency (Hari et al., 2020; Markonis et al., 2021), and accumulating scientific evidence highlights their negative impacts on ecosystems, especially on forests ecosystems (Bastos et al., 2020, 2021; Forzieri et al., 2021; Gampe et al., 2021). Several studies reported widespread premature leaf senescence in 2018, unprecedented drought-induced tree mortality across various species throughout the region and reductions in tree growth (Bose et al., 2020; Brun et al., 2020; Buras et al., 2020; Schnabel et al., 2022; Schuldt et al., 2020). Tree stress responses were found to be even more pronounced in 2019 than in 2018, indicating that consecutive and compound drought years represent a novel stressor for forests (Schnabel et al., 2022). Lags in physiological recovery, that is, drought legacy effects, can be caused by hydraulic damage (Anderegg et al., 2018; Kannenberg et al., 2019), carbon depletion or shifts in carbon allocation (e.g. towards rebuilding the canopy, growing roots or reproduction), which manifest in the reduction of radial stem growth.
For the quantification of drought impacts, the concept of resistance and resilience could be applied to forest communities to unravel these different facets of ecosystem stability (Ingrisch & Bahn, 2018; Isbell et al., 2015; Lloret et al., 2011). Although there are various approaches to address components of stability (for an overview, see Ingrisch & Bahn, 2018), we consider resistance here as the ability to persist and maintain functioning during a disturbance quantified as the ratio between tree growth during drought and tree growth during the respective pre-drought period, characterized by ‘normal’ climate conditions (Lloret et al., 2011). Similarly, resilience is defined as the capacity to reach pre-disturbance performance levels, and is estimated as the ratio between post-drought growth and pre-drought growth (Lloret et al., 2011).
Therefore, evaluating the differences in drought resistance and resilience among tree communities represents a crucial step towards understanding how the impacts of drought on forests could be mitigated by the choice of tree species and the design of climate-smart mixtures (Messier et al., 2022).
Biodiversity is known to stabilize ecosystem productivity over time and is therefore considered a key feature that supports the resistance and resilience of ecosystem functions to droughts (Cardinale et al., 2013; Isbell et al., 2015; Jourdan et al., 2020; Morin et al., 2014). In forest ecosystems, the influence of diversity on drought resistance and resilience is attributed to beneficial interactions among tree species such as resource partitioning (e.g. differential stomatal regulation strategies), facilitation (e.g. active hydraulic redistribution) or selection effects (e.g. competitive dominance of deep-rooted species) (Grossiord, 2020). Experimental evidence from subtropical tree communities suggests that the stabilizing effect of tree species richness is driven by interannual variations in the growth of different tree species, which buffer the community against stress-related growth declines (Schnabel et al., 2021).
However, it remains unknown whether this positive diversity effect persists when communities experience an unprecedented drought episode—such as the 2018–2020 drought. Indeed, there are indications that the positive diversity effects observed under moderate drought stress may shift to negative effects due to competitive species interactions (Haberstroh & Werner, 2022). Evidence on the impact of tree diversity on forest growth during and after drought remains inconsistent with positive, but also neutral, and negative diversity effects on tree responses to drought being reported (Forrester et al., 2016; Gillerot et al., 2021; Grossiord, 2020; Grossiord et al., 2014; Jucker et al., 2014; Pardos et al., 2021).
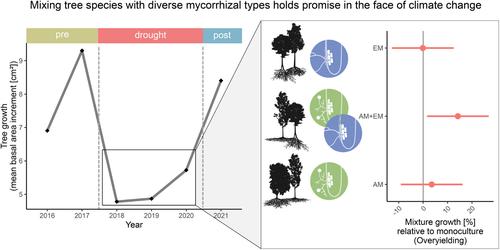
One possible factor explaining these inconsistent results may be the type of mycorrhizal association of the tree communities. Mycorrhizal fungi are known to help plants acquire nutrients and water uptake in exchange for photosynthates (Bowles et al., 2018; Lehto & Zwiazek, 2011), as mycorrhizal hyphae reach soil water and nutrients that would be inaccessible to plant roots (Allen, 2007). Therefore, the type of mycorrhizal association could play an important role in the effects of drought on forests. A growing body of research suggests that the type of mycorrhizal association is a key driver for ecosystem-functioning relationships (Deng et al., 2023; Luo et al., 2023; Mao et al., 2023). There are two main groups of mycorrhizal association types that are formed between temperate tree species and fungi: ectomycorrhiza (EM) and arbuscular mycorrhiza (AM), which differ in their morphology, physiology and therefore soil nutrient uptake processes (Phillips et al., 2013; Tedersoo & Bahram, 2019). Ectomycorrhizal fungi develop a mantle of hyphae around the tips of the plant roots through which the exchange of nutrients with their hosts occurs. Arbuscular mycorrhizal fungi are endophytic and exchange nutrients within the inner cortical cells of the plant host fine roots (Peterson & Massicotte, 2004). While AM fungi primarily provide their plant host with access to soil phosphorus in the upper mineral soil layer, EM fungi can mobilize both organic and mineral plant resources and typically thrive in organic soil horizons (Midgley & Phillips, 2014; Phillips et al., 2013; Read & Perez-Moreno, 2003; Rosling et al., 2016; Toju et al., 2016). The differences between mycorrhizal types in relation to the benefit provided to their host trees under drought conditions remain unclear, due to a lack of comparative studies (Allen, 2007; Gehring et al., 2006; Kilpeläinen et al., 2017; Mohan et al., 2014; Querejeta et al., 2009; Teste et al., 2020). Although dual mycorrhization with AM and EM in plant roots seems common (Teste et al., 2020), one of the two mycorrhizal types dominates the association in most temperate tree species (Ferlian et al., 2021; Heklau et al., 2021). Due to the distinct lifestyles and foraging strategies of AM and EM fungi, it can be expected that the presence of both types of association within a plant community could lead to higher resource partitioning among their associated plant hosts (Ferlian et al., 2018; Luo et al., 2018; Teste et al., 2020; Wagg, Jansa, Schmid, et al., 2011; Wagg, Jansa, Stadler, et al., 2011). Particularly during drought stress, when tree communities lack water and nutrient supply, the potential benefit of mycorrhizal type richness, which can offer support by complementary resource access (Ferlian et al., 2018; Teste et al., 2020), may become more pronounced.
However, evidence of the promising role of mycorrhizal associations in drought resistance and resilience of tree communities and especially studies using both types of mycorrhizal association are lacking so far. For the determination of management actions for forests in future climatic conditions, it is crucial to understand the relevance of both tree diversity and below-ground mycorrhizal associations, for forest resistance and resilience to drought (Eisenhauer et al., 2022). Such insights could be best achieved with an experimental approach, which manipulates both factors while controlling for confounding environmental effects (Eisenhauer et al., 2022; Ferlian et al., 2018; Scherer-Lorenzen et al., 2005).
Here, we evaluated the growth resistance and resilience of tree communities varying in tree species richness to the 2018–2020 drought, using inventory data from a tree diversity experiment in Germany (MyDiv) that crosses tree species richness (monocultures, 2-species mixtures, 4-species mixtures) with mycorrhizal association types (AM only, EM only and AM + EM).
Specifically, we tested the following hypotheses:
H1.Growth resistance and resilience to the 2018–2020 drought increase with tree species richness.
H2.Communities that contain tree species of both mycorrhizal association types (AM + EM) exhibit higher growth resistance and resilience to the 2018–2020 drought compared to tree communities with either AM associations or EM associations alone.
H3.The relationship between growth resistance and resilience to the 2018–2020 drought and tree species richness is modulated by the mycorrhizal association type of the tree communities.
中文翻译:
森林生长对 2018-2020 年干旱的抵抗力和恢复力取决于树木多样性和菌根类型
1 简介
全球变暖导致严重和连续干旱的可能性增加(Hari等人, 2020年;Spinoni等人, 2018年)。森林生态系统尤其面临干旱的威胁,因为与其他植物相比,树木的世代时间长且生长缓慢(Brodribb 等, 2020 )。这是一个令人高度关注的问题,因为除了保护生物多样性和其他生态系统服务外,森林还作为碳汇发挥着至关重要的作用,从而有助于缓解当前和未来的气候变化(Anderegg等人, 2020 )。
2018年,北欧和中欧经历了一场非同寻常的复合干旱,其特点是降水不足和热浪(Zscheischler等, 2020 ;Zscheischler&Fischer, 2020 )。异常干燥的土壤条件在 2019 年持续存在,在中欧广大地区,甚至持续到 2020 年(Rakovec 等, 2022 )。 2018年至2020年连续三年的干旱(以下简称“2018-2020年干旱”)的出现,标志着中欧至少在过去250年中经历了前所未有的干旱局面(Bastos等, 2021 ;Hari等人, 2020 ;Rakovec 等人, 2022 )。这种复合和连续干旱的频率大幅增加(Hari等人, 2020年;Markonis等人, 2021年),并且不断积累的科学证据强调了它们对生态系统,特别是森林生态系统的负面影响(Bastos等人, 2020年, 2021年) ;Forzieri 等人, 2021 ;Gampe 等人, 2021 )。多项研究报告称,2018 年,叶子普遍早衰,该地区各种物种因干旱而死亡,树木生长减少(Bose 等人, 2020 年;Brun 等人, 2020 年;Buras 等人, 2020 年;Schnabel)等人, 2022 ;舒尔特等人, 2020 )。研究发现,2019 年树木的应激反应比 2018 年更加明显,这表明连续和复合的干旱年份对森林来说是一种新的应激源(Schnabel 等,2018)。, 2022 )。生理恢复的滞后,即干旱遗留效应,可能是由水力损害(Anderegg等人, 2018年;Kannenberg等人, 2019年)、碳枯竭或碳分配的变化(例如重建树冠、生长根系)引起的或繁殖),这表现为径向茎生长的减少。
为了量化干旱影响,抵抗力和复原力的概念可以应用于森林群落,以揭示生态系统稳定性的不同方面(Ingrisch & Bahn, 2018 ;Isbell 等, 2015 ;Lloret 等, 2011 )。尽管有多种方法可以解决稳定性的组成部分(概述,请参见 Ingrisch & Bahn, 2018 ),但我们将此处的抵抗力视为在干扰期间持续和维持功能的能力,量化为干旱期间树木生长与树木生长之间的比率在相应的干旱前期,其特点是“正常”气候条件(Lloret 等人, 2011 年)。同样,恢复力被定义为达到扰动前绩效水平的能力,并被估计为干旱后增长与干旱前增长之间的比率(Lloret 等, 2011 )。
因此,评估树木群落抗旱性和复原力的差异是了解如何通过选择树种和设计气候智能型混合物来减轻干旱对森林的影响的关键一步(Messier et al., 2022) )。
众所周知,生物多样性可以随着时间的推移稳定生态系统生产力,因此被认为是支持生态系统功能抵御干旱和恢复能力的关键特征(Cardinale 等人, 2013 年;Isbell 等人, 2015 年;Jourdan 等人, 2020 年;Morin)等人, 2014 )。在森林生态系统中,多样性对抗旱性和复原力的影响归因于树种之间的有益相互作用,例如资源分配(例如差异性气孔调节策略)、便利化(例如主动水力再分配)或选择效应(例如深层树种的竞争优势)。有根物种)(Grossiord, 2020 )。来自亚热带树木群落的实验证据表明,树种丰富度的稳定作用是由不同树种生长的年际变化驱动的,这可以缓冲群落免受与压力相关的生长下降的影响(Schnabel et al., 2021 )。
然而,当社区经历前所未有的干旱事件(例如 2018 年至 2020 年的干旱)时,这种积极的多样性效应是否持续存在仍不得而知。事实上,有迹象表明,由于竞争性物种相互作用,在中度干旱胁迫下观察到的积极多样性效应可能会转变为负面效应(Haberstroh&Werner, 2022 )。关于干旱期间和干旱后树木多样性对森林生长影响的证据仍然与所报告的树木多样性对干旱反应的积极、中性和消极影响不一致(Forrester 等人, 2016 年;Gillerot 等人, 2021 年;Grossiord) , 2020 ;Grossiord 等, 2014 ;Jucker 等, 2014 ;Pardos 等, 2021 )。
解释这些不一致结果的一个可能因素可能是树木群落的菌根关联类型。众所周知,菌根真菌可以帮助植物获取养分和水分吸收,以换取光合产物(Bowles et al., 2018 ;Lehto & Zwiazek, 2011 ),因为菌根菌丝可以到达植物根部无法获取的土壤水分和养分(Allen, 2007) )。因此,菌根联合体的类型可能在干旱对森林的影响中发挥重要作用。越来越多的研究表明,菌根关联的类型是生态系统功能关系的关键驱动因素(Deng 等, 2023 ;Luo 等, 2023 ;Mao 等, 2023 )。温带树种和真菌之间形成的菌根关联类型主要有两类:外生菌根 (EM) 和丛枝菌根 (AM),它们的形态、生理学和土壤养分吸收过程不同(Phillips 等, 2013 )特德苏和巴赫拉姆, 2019 )。外生菌根真菌在植物根尖周围形成菌丝外套膜,通过菌丝外套膜与宿主进行营养物质交换。丛枝菌根真菌是内生真菌,在植物宿主细根的内皮层细胞内交换养分(Peterson & Massicotte, 2004 )。 AM 真菌主要为植物宿主提供上层矿质土壤层的土壤磷,而 EM 真菌可以调动有机和矿物植物资源,并且通常在有机土壤层中茁壮成长(Midgley & Phillips, 2014 ;Phillips 等,2014)。, 2013 ;里德和佩雷斯-莫雷诺, 2003 年;罗斯林等人, 2016 ; Toju 等人, 2016 )。由于缺乏比较研究,菌根类型之间的差异与其在干旱条件下为寄主树提供的益处之间的差异仍不清楚(Allen, 2007 ;Gehring 等, 2006 ;Kilpeläinen 等, 2017 ;Mohan 等) ., 2014 ;Querejeta 等人, 2009 ;Teste 等人, 2020 )。尽管植物根部 AM 和 EM 双重菌根化似乎很常见(Teste 等人, 2020 ),但这两种菌根类型中的一种在大多数温带树种中占主导地位(Ferlian 等人, 2021 ;Heklau 等人, 2021 ) 。由于 AM 和 EM 真菌不同的生活方式和觅食策略,可以预期,植物群落内两种类型关联的存在可能会导致其相关植物宿主之间更高的资源分配(Ferlian 等, 2018 ;Luo等人, 2018 ;Teste 等人, 2020 ;Wagg、Jansa、Schmid 等人, 2011 ;Wagg、Jansa、Stadler 等人, 2011 )。特别是在干旱胁迫期间,当树木群落缺乏水和养分供应时,菌根类型丰富度的潜在效益可能会变得更加明显,菌根类型丰富度可以通过补充资源获取提供支持(Ferlian et al., 2018 ;Teste et al., 2020 )。 。
然而,迄今为止,还缺乏菌根联合体在树木群落的抗旱性和恢复力方面的潜在作用的证据,特别是使用这两种类型的菌根联合体的研究。为了确定未来气候条件下的森林管理行动,了解树木多样性和地下菌根关联对于森林抵抗干旱和恢复力的相关性至关重要(Eisenhauer 等, 2022 )。这种见解最好通过实验方法来实现,该方法在控制混杂的环境影响的同时操纵这两个因素(Eisenhauer 等人, 2022 ;Ferlian 等人, 2018 ;Scherer-Lorenzen 等人, 2005 )。
在这里,我们使用德国一项跨越树种丰富度(单一栽培、2种混合、 4 种混合物)与菌根联合类型(仅 AM、仅 EM 和 AM + EM)。
具体来说,我们测试了以下假设:
H1。随着树种的丰富,生长对 2018-2020 年干旱的抵抗力和恢复力也会增强。
H2。与仅具有 AM 关联或 EM 关联的树群落相比,包含两种菌根关联类型 (AM + EM) 树种的群落对 2018-2020 年干旱表现出更高的生长抗性和恢复力。
H3。 2018-2020年干旱的生长抗性和恢复力与树种丰富度之间的关系受到树木群落菌根关联类型的调节。






























 京公网安备 11010802027423号
京公网安备 11010802027423号