Journal of Ecology ( IF 5.3 ) Pub Date : 2024-07-08 , DOI: 10.1111/1365-2745.14367 Insu Jo 1 , Peter J. Bellingham 1 , Norman W. H. Mason 2 , James K. McCarthy 1 , Duane A. Peltzer 1 , Sarah J. Richardson 1 , Elaine F. Wright 3
|
1 INTRODUCTION
Disturbance, whether natural or anthropogenic, can promote establishment of non-native plants by creating open space with little competition from existing plants or increased resource availability (Davis et al., 2000; Hobbs & Huenneke, 1992). For example, flooding and moderate fire disturbances can facilitate invasion by non-native plant species by removing the resident vegetation and increasing soil nutrient availability through alteration of below-ground microbial communities or organic substrate structure (Certini, 2005; Frey et al., 2003; Molles Jr et al., 1998). Similarly, many empirical studies report greater invader abundance and richness with increasing disturbance intensity (Alston & Richardson, 2006; Lake & Leishman, 2004; Lembrechts et al., 2016; Parks et al., 2005; Seabloom et al., 2003), particularly where disturbance increases nutrient availability (Lake & Leishman, 2004; Lembrechts et al., 2016). However, most of these studies focus on local contexts (Daniels & Larson, 2020; Eschtruth & Battles, 2009); what remains poorly understood is whether the multiple mechanisms underpinning disturbance-mediated plant invasions also occur at larger spatial scales in natural ecosystems, particularly in forests. Studies quantifying invasion across wide environmental gradients that capture the wide suite of variables influenced by disturbances such as structure and functional composition of resident communities, soil fertility, and the functional strategies of invading species can provide deeper insights into the mechanisms of disturbance-driven invasions in natural ecosystems, (Rossignaud & Hulme, 2023; Stohlgren et al., 2003).
Forest understories are widely thought to be more resistant to non-native plant invasions than other habitats due to low light availability (Catford et al., 2012; Nunez-Mir et al., 2017), but could also reflect lower propagule pressure or greater invasion lags in forests (Essl et al., 2012). Forests are complex, dynamic systems characterised by different small- and large-scale disturbances which can significantly alter canopy properties, species composition and understorey structure (Attiwill, 1994). For example, mortality of large trees creates canopy gaps that increase light availability at the ground level and allowing a window of opportunity for species recruitment or regeneration (Canham et al., 1990). Landslides, fire, and logging can remove resident vegetation, promoting recruitment of non-native species into newly opened areas (Restrepo & Vitousek, 2001; Richardson, 1988). Shifts in forest composition following disturbance can also alter nutrient cycling through multiple mechanisms including shifts in plant functional traits or types (e.g. ectomycorrhizal vs. arbuscular mycorrhizal, Phillips et al., 2013). Biotic disturbances like herbivory also affect understorey vegetation through selective consumption of palatable species (Averill et al., 2018; Côté et al., 2004; Gill, 1992) and these interactions are strongly linked to nutrient cycling and other ecosystem processes (Bardgett & Wardle, 2003; Schmitz et al., 2018; Schmitz & Leroux, 2020). Furthermore, herbivores can disperse seeds of native and non-native species and add nutrients (via urine and faeces), leading to increases in soil nutrient availability (Liu et al., 2018; Vavra et al., 2007). What this literature suggests is that multiple disturbance-mediated changes in understorey resource availability often occur, but whether these processes facilitate establishment and proliferation of non-native plant species in forests at larger spatial scales is rarely evaluated (Davis et al., 2000; Martin et al., 2008; Wiser et al., 1998).
Invasive plant species often have different functional strategies than co-occurring native species (Jo et al., 2015; Ordonez et al., 2010). Meta-analyses comparing multiple functional traits of native and non-native invasive plant species show that non-native invaders often have traits associated with greater growth and reproduction rates relative to native species (Leishman et al., 2007; van Kleunen et al., 2010). However, invaders in forest understories may not necessarily exhibit the typical traits associated with faster growth and higher reproduction rates. For example, in undisturbed forest understories where light availability is low, tall-stature species (e.g. woody plants) have a greater ability to capture light than herbaceous species. This allows them to outcompete herbaceous species and dominate the understorey (Bazzaz, 1968). A recent study of understorey woody invaders supports this view; many non-native species can persist in forest understories at low light levels and then grow rapidly when light levels increase, following disturbance (Fridley et al., 2023). High propagule pressure (e.g. proximity to human-altered landscapes or source populations) can also drive invasion success, even for species lacking the functional traits associated with high performance (Davis et al., 2000; Ni et al., 2021). Despite the rich literature describing biological invasions, the influence of disturbances and disturbance-mediated community characteristics on the functional strategies of co-occurring native and non-native species has rarely been quantified or investigated across broad environmental gradients and varied forest communities (Perkins et al., 2011; Richardson & Pyšek, 2006).
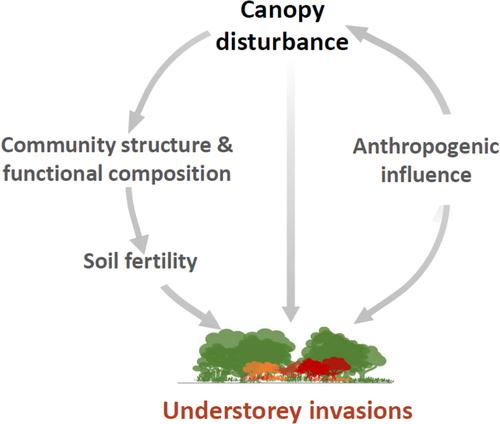
In this study, we aimed to disentangle the mechanisms underpinning disturbance-mediated invasions in natural forests, using systematically sampled forest vegetation inventory data throughout New Zealand's (NZ) main islands (34–47° S, 166–178° E), which cover a diverse climatic and soil fertility spectrum. In addition, there is a relatively large pool of non-native plant species (>1800 species) that are naturalised, many of which could potentially spread into native forest vegetation (Brandt et al., 2021). First, we determined how anthropogenic influence is related to canopy disturbance (Q1) and whether disturbance increases non-native plant species invasions in forest understories (Q2). Then, we tested how disturbance-mediated changes in forest community structure and functional composition are associated with soil fertility and wild ungulate abundance, and how these disturbance-mediated community characteristics affect understorey invasions (Q3). Finally, we determined whether functional assemblages of co-occurring native and non-native species differ depending on plot disturbance status (Q4) and how the functional assemblages of native and non-native plants respond differently to the disturbance-mediated invasion predictors (Q5).
The natural evergreen rain forests of NZ encompass a diverse range of ecosystems, from coastal to subalpine, spanning warm temperate to cold temperate climates (Newnham et al., 1999; Wardle, 1991). These forests include both old-growth communities and secondary communities (Bellingham et al., 1999; Holdaway et al., 2017; Richardson et al., 2009) that have regenerated following anthropogenic disturbances, such as fire and logging, or natural disturbances, such as landslides, earthquakes, and wind (Coomes & Allen, 2007; Dawson & Sneddon, 1969; Martin & Ogden, 2006; McGlone et al., 1996; Perry et al., 2014). Further, NZ has a large number of invasive non-native plant species and a good record of their introduction history (Brandt et al., 2021; Hulme, 2020; Timmins & Williams, 1991; Wilton & Breitwieser, 2000). These characteristics of NZ's forests and invasion history provide crucial context for investigating the mechanisms by which disturbance contributes to understorey invasions in natural forests.
中文翻译:
干扰介导的群落特征和人为压力加剧了天然林中的林下植物入侵
1 简介
干扰,无论是自然的还是人为的,都可以通过创造与现有植物几乎没有竞争的开放空间或增加资源可用性来促进非本地植物的建立(Davis 等, 2000 ;Hobbs 和 Huenneke, 1992 )。例如,洪水和适度的火灾扰动可以通过去除常驻植被并通过改变地下微生物群落或有机基质结构来增加土壤养分可用性,从而促进非本地植物物种的入侵(Certini, 2005 ;Frey等人, 2003 )小莫勒斯等人, 1998 )。同样,许多实证研究报告称,随着干扰强度的增加,入侵者的丰度和丰富度也随之增加(Alston & Richardson, 2006 ;Lake & Leishman, 2004 ;Lembrechts 等, 2016 ;Parks 等, 2005 ;Seabloom 等, 2003 ),特别是在干扰增加养分可用性的情况下(Lake & Leishman, 2004 ;Lembrechts 等, 2016 )。然而,这些研究大多数都集中在当地背景(Daniels & Larson, 2020 ;Eschtruth & Battles, 2009 );人们仍然知之甚少的是,支持干扰介导的植物入侵的多种机制是否也发生在更大的空间尺度的自然生态系统中,特别是在森林中。 对广泛环境梯度中的入侵进行量化研究,捕捉受干扰影响的一系列变量,例如居民群落的结构和功能组成、土壤肥力以及入侵物种的功能策略,可以更深入地了解干扰驱动的入侵机制。自然生态系统,(Rossignaud & Hulme, 2023 ;Stohlgren 等人, 2003 )。
人们普遍认为,由于光照不足,森林下层植物比其他栖息地更能抵抗非本地植物入侵(Catford 等人, 2012 年;Nunez-Mir 等人, 2017 年),但也可能反映了较低的繁殖压力或较高的繁殖压力。森林的入侵滞后(Essl 等, 2012 )。森林是复杂的动态系统,其特征是不同的小规模和大规模干扰,可以显着改变树冠特性、物种组成和林下结构(Attiwill, 1994 )。例如,大树的死亡会产生树冠间隙,从而增加地面的光照可用性,并为物种补充或再生提供机会(Canham 等, 1990 )。山体滑坡、火灾和伐木可以清除常驻植被,促进非本地物种进入新开放区域(Restrepo & Vitousek, 2001 ;Richardson, 1988 )。干扰后森林组成的变化也可以通过多种机制改变养分循环,包括植物功能性状或类型的变化(例如外生菌根与丛枝菌根,Phillips等, 2013 )。食草等生物干扰也会通过选择性消耗适口物种来影响林下植被(Averill 等, 2018 ;Côté 等, 2004 ;Gill, 1992 ),这些相互作用与养分循环和其他生态系统过程密切相关(Bardgett 和 Wardle) , 2003 ;施密茨等人, 2018 ;施密茨和勒鲁, 2020 )。 此外,食草动物可以传播本地和非本地物种的种子并添加养分(通过尿液和粪便),从而增加土壤养分的可用性(Liu et al., 2018 ;Vavra et al., 2007 )。该文献表明,林下资源可用性经常发生多种干扰介导的变化,但这些过程是否促进更大空间尺度的森林中非本地植物物种的建立和增殖却很少被评估(Davis 等, 2000 ;Martin等人, 2008 ;Wiser 等人, 1998 )。
入侵植物物种通常具有与共存本地物种不同的功能策略(Jo 等人, 2015 年;Ordonez 等人, 2010 年)。比较本地和非本地入侵植物物种的多种功能性状的荟萃分析表明,与本地物种相比,非本地入侵者通常具有与更高的生长和繁殖率相关的特征(Leishman 等人, 2007 年;van Kleunen 等人, 2010 )。然而,森林林下的入侵者不一定表现出与更快的生长和更高的繁殖率相关的典型特征。例如,在光利用率较低的未受干扰的森林林下,高大物种(例如木本植物)比草本物种捕获光的能力更强。这使得它们能够在竞争中击败草本物种并主宰林下植被(Bazzaz, 1968 )。最近一项针对林下木质入侵者的研究支持了这一观点。许多非本地物种可以在低光照水平下持续存在于森林林下,然后在受到干扰后,当光照水平增加时迅速生长(Fridley et al., 2023 )。高繁殖压力(例如接近人类改变的景观或来源种群)也可以推动入侵成功,即使对于缺乏与高性能相关的功能特征的物种也是如此(Davis 等人, 2000 年;Ni 等人, 2021 年)。 尽管有丰富的文献描述生物入侵,但干扰和干扰介导的群落特征对共存的本地和非本地物种的功能策略的影响很少在广泛的环境梯度和不同的森林群落中进行量化或研究(Perkins等人) ., 2011 ;理查森和皮谢克, 2006 )。
在本研究中,我们旨在利用新西兰 (NZ) 主要岛屿(南纬 34–47°,东经 166–178°)系统采样的森林植被清查数据,阐明天然林中干扰介导的入侵的机制,这些数据涵盖多样化的气候和土壤肥力范围。此外,还有相对较大的非本地植物物种(> 1800 种)已归化,其中许多有可能扩散到本地森林植被中(Brandt 等, 2021 )。首先,我们确定了人为影响与冠层干扰之间的关系(Q1)以及干扰是否会增加森林下层非本地植物物种的入侵(Q2)。然后,我们测试了干扰介导的森林群落结构和功能组成的变化如何与土壤肥力和野生有蹄类动物丰度相关,以及这些干扰介导的群落特征如何影响林下入侵(Q3)。最后,我们确定了共存的本地和非本地物种的功能组合是否因地块干扰状态而不同(Q4),以及本地和非本地植物的功能组合如何对干扰介导的入侵预测因子做出不同的反应(Q5) 。
新西兰的天然常绿雨林涵盖了多种生态系统,从沿海到亚高山,跨越暖温带到冷温带气候(Newnham 等, 1999 ;Wardle, 1991 )。这些森林包括原始群落和次生群落(Bellingham 等, 1999 ;Holdaway 等, 2017 ;Richardson 等, 2009 ),它们在人为干扰(例如火灾和伐木)或自然干扰后再生,例如山体滑坡、地震和风(Coomes & Allen, 2007 ;Dawson & Sneddon, 1969 ;Martin & Ogden, 2006 ;McGlone et al., 1996 ;Perry et al., 2014 )。此外,新西兰拥有大量入侵性非本地植物物种,并且对其引入历史有良好的记录(Brandt et al., 2021 ; Hulme, 2020 ; Timmins & Williams, 1991 ; Wilton & Breitwieser, 2000 )。新西兰森林的这些特征和入侵历史为研究干扰导致天然林林下入侵的机制提供了重要的背景。






























 京公网安备 11010802027423号
京公网安备 11010802027423号