Journal of Ecology ( IF 5.3 ) Pub Date : 2024-04-03 , DOI: 10.1111/1365-2745.14306 Tyson J. Terry 1, 2 , Peter B. Adler 2
|
1 INTRODUCTION
Ecological disturbances, natural or anthropogenic, cause long-lasting changes to vegetation in dryland systems (Allred et al., 2015; Boyd & Davies, 2012) and have been linked to shifts in stable states (Abella et al., 2021). Understanding recovery trajectories following disturbance is necessary for guiding management and conservation of lands that are susceptible to long-term impacts (Chambers et al., 2017). However, great uncertainty exists concerning recovery rates and how they depend on environmental factors in drylands (Stafford Smith et al., 2009). This uncertainty reflects the short-term nature or limited spatial scale of previous disturbance studies (O'Brien et al., 2022).
The lack of spatially extensive disturbance studies in drylands stems from multiple challenges that make it difficult to compare impacts of different disturbances occurring in separate locations. Disturbances vary in type, severity, timing and patchiness, all of which influence post-disturbance dynamics and complicate spatial comparisons (Bartels et al., 2016; Díaz-Delgado et al., 2003). Furthermore, vegetation types characterized by different disturbance regimes usually have different climates. Funding generally limits the spatial and temporal scale of recovery monitoring such that disturbance data is often post hoc, comprised of many space-for-time substitutions that may not capture long-term recovery pathways. Many studies compare the value of a response variable after the disturbance to a pre-disturbance baseline, but this approach may also be unreliable because it assumes a steady-state equilibrium that is unlikely in an era of changing climate and invasive species (Monroe et al., 2022; Parker & Wiens, 2005).
Recovery, which we define as the return of a variable to its undisturbed state following a perturbation (Chambers et al., 2019; Oliver et al., 2015), comprises two processes: the initial, short-term response of a variable to the disturbance and subsequent changes over time following the initial response (Hodgson et al., 2015; Ingrisch & Bahn, 2018). These two processes are comparable to resistance (degree of initial response) and engineering resilience (rate of recovery after initial response) (Ingrisch & Bahn, 2018; Nimmo et al., 2015).
A common hypothesis is that the rate of recovery of plant communities following a disturbance depends on resource availability (Chapin et al., 1996; Shriver et al., 2018; Tilman, 2016). In drylands, water availability is the key resource driving net primary production (NPP) and vegetation dynamics (Chambers et al., 2014; Jordan et al., 2020), with previous studies showing that wetter areas or weather periods promote recruitment success (Nelson et al., 2014; O'Connor et al., 2020). However, recovery of a system following disturbance requires a return to current undisturbed levels of function and structure (Chambers et al., 2019; Oliver et al., 2015), and many of the studies supporting this hypothesis in drylands do not compare post-disturbance growth with contemporary undisturbed controls.
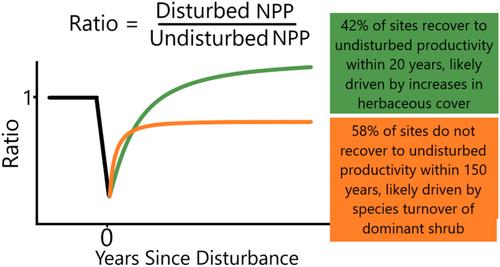
An alternative hypothesis we propose, is that recovery in drylands may be faster in low-resource environments with sparse vegetation. For example, dry areas might recover more quickly than wet areas due to the small amount of growth needed to recover to the relatively low level of productivity found in dry undisturbed control sites, whereas wetter areas may require substantially more time to recover the comparatively high levels of productivity found in wetter undisturbed sites. This relative-recovery hypothesis emphasizes the importance of undisturbed reference conditions to quantify recovery. Moreover, including undisturbed controls becomes necessary to compare recovery across climatic gradients (Parker & Wiens, 2005), especially in drylands where primary production potential and species composition are strongly tied to water availability (Sala et al., 2012).
Recovery from disturbance often depends on recruitment, which in drylands is tightly linked with soil water availability (Bradford et al., 2019; Nelson et al., 2014). Water availability in drylands is largely determined by interactions between precipitation quantity, seasonality and soil texture (Loik et al., 2004; Renne et al., 2019). Coarse soil texture may benefit deep-rooted plants in arid conditions, where high infiltration reduces loss of soil water to evaporation and increases water availability in deep soil layers (Maurer et al., 2020; Noy-Meir, 1973; Walter, 1964). However, the benefits of coarse soil texture decline with increases in total precipitation, as losses of soil water to deep drainage outweigh the benefits of low evaporative losses (Noy-Meir, 1973). This idea, commonly known as the inverse texture hypothesis, is largely supported by studies of NPP (Sala et al., 1988) and mature plant abundance (Renne et al., 2019). However, the effect of coarse soils on recruitment remains unclear for shallow-rooted seedlings (Barnard et al., 2019; Boyd & Davies, 2012). Seasonal timing of precipitation also impacts evaporative losses (Lauenroth & Bradford, 2012) that are thought to determine the benefits of coarse soil texture for plant growth (Renne et al., 2019). Testing hypotheses about water availability and recovery following disturbance in drylands therefore requires careful consideration of interactions between seasonal precipitation regimes and soil properties.
Natural gas pipeline corridors provide an opportunity to investigate variation in recovery across environmental gradients, overcoming common limitations of traditional disturbance studies. Pipeline corridors create a near-uniform pulse disturbance that runs hundreds of kilometres, spanning broad soil and climate gradients. Pipeline installation consists of removing all above-ground biomass via bulldozer in a strip we refer to as the corridor (up to 35 m wide), followed by digging a trench and burying a 20- to 30-cm-diameter pipe down the centre of the cleared corridor. This disturbance not only removes all plants but also displaces and compacts surface soil (Shi et al., 2014). Following construction, topsoils are spread back over the corridor and then seeded for restoration. Post-construction seeding efforts generally use seed mixes to match native species and functional types, but these efforts are often unsuccessful and with mixed effects on species composition (Farrell & Fehmi, 2018; Rottler et al., 2018). Construction effects are largely concentrated within the pipeline corridors, leaving undisturbed neighbouring vegetation and soils as a control to measure recovery. Instead of relying on historical, pre-disturbance conditions as the baseline for measuring recovery, this data set allows yearly comparisons of disturbed and undisturbed vegetation to account for changes over time in undisturbed sites.
Here, we studied two dimensions of ecosystem recovery following pipeline disturbance: total shrub cover and NPP. Shrub cover represents a dominant plant functional type in North American drylands (Peinado et al., 1995), where many imperilled wildlife species are considered shrub obligates (Suring et al., 2005). NPP represents the rate at which energy enters the ecosystem and is an indicator of dryland degradation (Wessels et al., 2008; Zika & Erb, 2009). We used annual remotely sensed estimates of NPP and total shrub cover along natural gas pipeline corridors to answer two research questions: (1) How long does it take for NPP and shrub cover in drylands to recover following a disturbance that removes all biomass and disrupts the surface soil? (2) How do mean annual precipitation (MAP), precipitation seasonality and soil texture interact to influence time to recovery of NPP and shrub cover in drylands? We hypothesized that interactions between climate and soils that increase water availability could either (a) increase recruitment and speed up recovery of both NPP and shrub cover, or (b) create high undisturbed values of NPP and shrub cover that require more time to recover.
中文翻译:
降水季节性和土壤质地相互作用,影响旱地从严重扰动中的恢复
1 简介
自然或人为的生态干扰会导致旱地系统植被发生长期变化(Allred 等,2015;Boyd & Davies,2012),并与稳定状态的变化有关(Abella 等,2021)。了解干扰后的恢复轨迹对于指导易受长期影响的土地的管理和保护是必要的(Chambers 等,2017)。然而,旱地的恢复率及其对环境因素的依赖程度存在很大的不确定性(Stafford Smith 等,2009)。这种不确定性反映了先前扰动研究的短期性质或有限的空间规模(O'Brien 等人,2022)。
旱地缺乏广泛的空间扰动研究源于多重挑战,这些挑战使得很难比较不同地点发生的不同扰动的影响。扰动的类型、严重程度、时间和斑块性各不相同,所有这些都会影响扰动后的动态并使空间比较变得复杂(Bartels 等人,2016 年;Díaz-Delgado 等人,2003 年)。此外,以不同干扰方式为特征的植被类型通常具有不同的气候。资金通常会限制恢复监测的空间和时间规模,因此扰动数据通常是事后的,由许多可能无法捕获长期恢复路径的时空替代组成。许多研究将干扰后的响应变量值与干扰前的基线进行比较,但这种方法也可能不可靠,因为它假设了稳态平衡,这在气候变化和入侵物种的时代是不可能的(Monroe等人) .,2022;帕克和维恩斯,2005)。
恢复,我们将其定义为变量在扰动后恢复到未受扰动的状态(Chambers et al., 2019; Oliver et al., 2015),包括两个过程:变量对扰动的初始、短期响应。初始响应后随时间的干扰和后续变化(Hodgson 等人,2015 年;Ingrisch & Bahn,2018 年)。这两个过程相当于阻力(初始响应程度)和工程弹性(初始响应后的恢复率)(Ingrisch & Bahn,2018;Nimmo 等,2015)。
一个常见的假设是,植物群落在干扰后的恢复速度取决于资源的可用性(Chapin 等,1996;Shriver 等,2018;Tilman,2016)。在旱地,水资源供应是驱动净初级生产力(NPP)和植被动态的关键资源(Chambers 等,2014;Jordan 等,2020),之前的研究表明,湿润地区或天气时期促进补充成功(Nelson等人,2014;奥康纳等人,2020)。然而,扰动后系统的恢复需要恢复到当前未受扰动的功能和结构水平(Chambers et al., 2019; Oliver et al., 2015),并且许多支持干旱地区这一假设的研究并没有比较事后的情况。通过当代不受干扰的控制来抑制干扰的增长。
我们提出的另一个假设是,在植被稀疏的资源匮乏环境中,旱地的恢复可能会更快。例如,干燥地区可能比潮湿地区恢复得更快,因为在干燥未受干扰的对照地点恢复到相对较低的生产力水平所需的生长量较小,而湿润地区可能需要更多的时间才能恢复相对较高的水平在潮湿、未受干扰的地点发现的生产力。这种相对恢复假说强调了未受干扰的参考条件对量化恢复的重要性。此外,为了比较不同气候梯度的恢复情况,有必要纳入未受干扰的控制(Parker & Wiens, 2005),特别是在初级生产潜力和物种组成与可用水量密切相关的旱地(Sala et al., 2012)。
从干扰中恢复通常取决于补充,这在旱地与土壤水分可用性密切相关(Bradford 等人,2019 年;Nelson 等人,2014 年)。旱地的可用水量很大程度上取决于降水量、季节性和土壤质地之间的相互作用(Loik 等,2004;Renne 等,2019)。粗糙的土壤质地可能有利于干旱条件下的深根植物,其中高渗透性减少了土壤水分蒸发损失,并增加了深层土壤的可用水量(Maurer 等,2020;Noy-Meir,1973;Walter,1964)。然而,粗土壤质地的好处随着总降水量的增加而下降,因为深层排水的土壤水损失超过了低蒸发损失的好处(Noy-Meir,1973)。这种想法通常被称为逆纹理假说,在很大程度上得到了 NPP(Sala 等人,1988)和成熟植物丰度(Renne 等人,2019)研究的支持。然而,粗糙土壤对浅根幼苗补充的影响仍不清楚(Barnard et al., 2019;Boyd & Davies, 2012)。季节性降水时间也会影响蒸发损失(Lauenroth & Bradford,2012),蒸发损失被认为决定了粗土壤质地对植物生长的益处(Renne 等人,2019)。因此,测试有关旱地干扰后水的可用性和恢复的假设需要仔细考虑季节性降水状况和土壤特性之间的相互作用。
天然气管道走廊提供了研究跨环境梯度恢复变化的机会,克服了传统扰动研究的常见局限性。管道走廊产生近乎均匀的脉冲扰动,绵延数百公里,跨越广阔的土壤和气候梯度。管道安装包括通过推土机清除我们称为走廊(宽达 35 m)的地带中的所有地上生物量,然后挖一条沟渠并在走廊中心埋入一根直径 20 至 30 厘米的管道。清理过的走廊。这种扰动不仅会清除所有植物,还会移动并压实表层土壤(Shi et al., 2014)。施工结束后,表土被铺回走廊上,然后播种以进行恢复。施工后播种工作通常使用种子混合物来匹配本地物种和功能类型,但这些努力往往不成功,并且对物种组成产生混合影响(Farrell & Fehmi,2018;Rottler 等,2018)。施工影响主要集中在管道走廊内,留下未受干扰的邻近植被和土壤作为衡量恢复情况的对照。该数据集不依赖历史的扰动前条件作为测量恢复的基线,而是允许对受扰动和未扰动的植被进行年度比较,以解释未扰动地点随时间的变化。
在这里,我们研究了管道干扰后生态系统恢复的两个维度:灌木丛总覆盖度和NPP。灌木覆盖代表了北美旱地的主要植物功能类型(Peinado 等,1995),其中许多濒临灭绝的野生动物物种被认为是灌木(Suring 等,2005)。 NPP 代表能量进入生态系统的速率,是旱地退化的指标(Wessels 等,2008;Zika 和 Erb,2009)。我们利用对天然气管道走廊沿线的 NPP 和灌木丛覆盖率的年度遥感估算来回答两个研究问题:(1)在一场消除了所有生物量并扰乱了干旱地区的 NPP 和灌木丛覆盖率需要多长时间才能恢复。表土? (2) 年平均降水量 (MAP)、降水季节和土壤质地如何相互作用影响旱地 NPP 和灌木覆盖的恢复时间?我们假设气候和土壤之间的相互作用增加了可用水量,可以(a)增加补充并加速 NPP 和灌木覆盖的恢复,或者(b)创造 NPP 和灌木覆盖的高未扰动值,需要更多的时间来恢复。
































 京公网安备 11010802027423号
京公网安备 11010802027423号