为了理解和预测植物-土壤微生物相互作用在植物竞争和共存中的作用,我们必须超越现象学描述,并考虑驱动反馈的潜在植物-土壤微生物相互作用。明确考虑微生物功能的机制框架将帮助我们预测微生物在不断变化的生物和非生物环境中对植物共存的影响,并确定它们相对于其他共存机制的重要性(Abbott 等,2021;Semchenko 等,2022 ; van der Putten等, 2016)。虽然微生物的功能极其多样化,但它们主要作为互利共生者、病原体或分解者与植物相互作用。在植物共存的背景下,PSF 研究主要集中在病原体上,因为强调负密度和频率依赖性。然而,有强有力的证据表明,共存促进反馈也可以通过共生体(例如固氮细菌和菌根真菌)和分解者来促进(例如Bennett等人,2017年;Bever,2002年;Siefert等人 , 2018 年; Veen)等人, 2019;惠特克等人, 2017)。打开土壤微生物黑匣子的机械方法可能比传统的现象学方法需要更多的测量(见第 6 节),但对于预测快速环境变化的后果至关重要,而无需在新环境中重复实验。我们的目标是综合关于调节植物之间资源竞争的微生物机制的最新证据,并提出一个基于资源的机制框架,用于理解和预测土壤微生物在植物竞争和共存中的作用(图1)。
图1
在图查看器中打开微软幻灯片软件
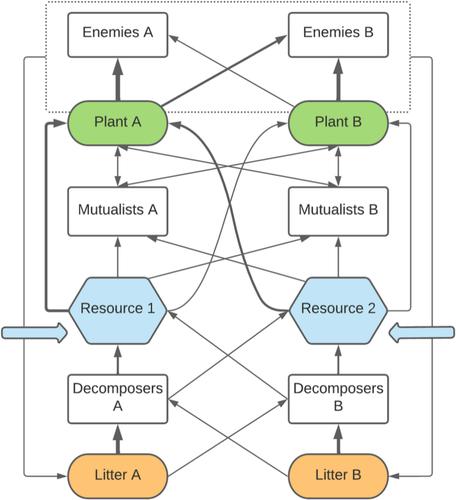
该概念图将不同的植物-土壤微生物相互作用纳入具有两个植物竞争者和两种资源的传统资源划分框架中。箭头代表能量和物质的流动,而不是效果。箭头的宽度表示流动的强度,反映了植物、微生物和资源之间假设的相互作用特异性(例如,这里每种植物更多地受到其特定敌人的攻击,并且其特定分解者处理更多宿主垃圾)。虚线框代表植物-敌害系统,该系统通过敌害引起的植物死亡而造成垃圾。蓝色箭头表示外部资源供应。当微生物缺乏宿主特异性时,可以通过合并每个功能组的微生物 A 和 B 成分来简化该图。
"点击查看英文标题和摘要"
Mechanistic approaches to investigate soil microbe-mediated plant competition
1 SOIL MICROBES MEDIATE PLANT COMPETITION
Plant–soil microbe interactions are ubiquitous, and recognition of their role in mediating plant competition has advanced our understanding of plant coexistence (Bever, 2003; Chung & Rudgers, 2016; Wagg et al., 2011). In the past few decades, negative feedback between plant hosts and their specific soil microbes (negative plant–soil feedback [PSFs]) have become the dominant paradigm for understanding the role of soil microbes in maintaining plant diversity (Crawford et al., 2019; Ke & Miki, 2015). This paradigm holds that plant species cultivate soil legacies via species-specific microbial assemblages that are more detrimental to themselves than co-occurring species; this self-limitation promotes coexistence.
Classic theory for PSFs is an extension of phenomenological models in which coexistence depends on the relative strength of intraspecific and interspecific interactions, without explicit reference to the mechanisms underlying those interactions (Bever et al., 1997; Revilla et al., 2013). This approach is experimentally tractable. The soil community can be treated as a single entity and evaluated based on its net effects on plant hosts. However, this tractability comes at the expense of mechanistic understanding. Recent reviews of PSFs have emphasized their importance (Gundale & Kardol, 2021), but warned of high context-dependence and low predictability (Bennett & Klironomos, 2019; De Long et al., 2019; Smith-Ramesh & Reynolds, 2017). For example, PSF magnitude and direction can change depending on plant age and development stage, resource availability, abiotic environmental gradients and host competitive context (Beals et al., 2020; Chung, 2023). We should not be satisfied with demonstrations that PSFs occur but should strive to quantify their contribution to the maintenance of diversity in nature.
To understand and predict the role of plant–soil microbe interactions in plant competition and coexistence, we must go beyond phenomenological description and consider the underlying plant–soil microbe interactions that drive the feedback. A mechanistic framework that explicitly considers microbial functions will help us predict microbial effects on plant coexistence in changing biotic and abiotic contexts and determine their importance relative to other coexistence mechanisms (Abbott et al., 2021; Semchenko et al., 2022; van der Putten et al., 2016). While microbial functions are hyperdiverse, they interact with plants primarily as mutualists, pathogens or decomposers. Within the context of plant coexistence, PSF research has focused primarily on pathogens due to the emphasis on negative density and frequency dependence. However, strong evidence exists that coexistence-promoting feedback can also be facilitated by mutualists, such as nitrogen-fixing bacteria and mycorrhizal fungi, and decomposers (e.g. Bennett et al., 2017; Bever, 2002; Siefert et al., 2018; Veen et al., 2019; Whitaker et al., 2017). Mechanistic approaches that open up the soil microbial black box may require more measurements than traditional phenomenological approaches (see Section 6) but are essential for predicting the consequences of rapid environmental change without the need to repeat experiments in new environments. We aim to synthesize recent evidence for microbial mechanisms that mediate resource competition between plants and propose a resource-based mechanistic framework for understanding and predicting the role of soil microbes in plant competition and coexistence (Figure 1).
FIGURE 1
Open in figure viewerPowerPoint
This conceptual diagram incorporates different plant–soil microbe interactions into a traditional resource partitioning framework with two plant competitors and two resources. Arrows represent the flow of energy and matter, not effects. The width of arrows indicates the strength of flow, which reflect hypothetical interaction specificity between plants, microbes, and resources (e.g. here each plant is attacked more by its specific enemy, and its specific decomposer processes more host litter). The dashed box represents the plant-enemy system, which contributes to litter via enemy-induced plant mortality. Blue arrows show external supply of resources. When microbes lack host specificity, this figure can be simplified by merging the microbial A and B components for each functional group.































 京公网安备 11010802027423号
京公网安备 11010802027423号