Insect Conservation and Diversity ( IF 3.2 ) Pub Date : 2023-06-23 , DOI: 10.1111/icad.12663 Pagi S. Toko 1, 2 , Bonny Koane 3 , Kenneth Molem 3 , Scott E. Miller 4 , Vojtech Novotny 1, 2
|
INTRODUCTION
Global biodiversity maxima, which include, for example, all six of the world's floristically richest areas hosting >5000 plant species per 10,000 km2 (Kier et al., 2005), often include long elevation gradients in the humid tropics, particularly the ‘complete’ rainforest elevation gradients that extend from lowlands to treeline, typically at ~3700 m asl. These gradients encompass a series of highly diverse rainforest ecosystems within a small geographic area, resulting from high species turnover along steep abiotic and biotic ecological gradients. These transects are ideal models for studying the mechanisms of community assembly without complicating effects of dispersal limitation. Recently, they have also become important in climate change research because they reproduce a wide range of temperatures in a limited area.
To a first approximation, species diversity shows either a monotonic decrease with increasing elevation, or a maximum at intermediate elevations (Rahbek, 1995; Stevens, 1992). The monotonic decline in species diversity may be directly or indirectly due to decreasing temperature and/or land area with increasing elevation (Beck & Chey, 2008; Beck & Kitching, 2009). Other important factors include primary productivity and habitat complexity, including vegetation structure important for birds (Sam et al., 2019), or the amount of litter important for ants (Moses et al., 2021; Pérez-Toledo et al., 2021). The maximum diversity at mid-elevation rarely corresponds to a single abiotic or biotic driving variable. It may be a product of the mid-domain effect (Colwell et al., 2016) and/or a combination of multiple factors. In particular, biotic pressures from predators and pathogens tend to decrease and abiotic stress tends to increase with increasing elevation (Péré et al., 2013). Unfortunately, trends in the intensity of trophic interactions such as herbivory, parasitism, or predation are particularly poorly known because they are often caused by multiple taxa that are rarely studied together (Bärtschi et al., 2019; Peters et al., 2016).
Gamma diversity along an entire elevation gradient is a product of alpha diversity values in individual communities and the rate of species turnover across elevations. The Rapoport rule suggests that beta diversity should decrease with increasing elevation, as montane species should have larger elevational ranges than lowland species (Stevens, 1992). However, trends in beta diversity along elevational gradients are studied less often than in alpha diversity (Grytnes & McCain, 2007). Beta diversity along an elevational gradient determines the overall extent of regional, transect-long gamma diversity. For example, a complete rainforest elevational gradient from lowland rainforest to treeline at Mt. Wilhelm in New Guinea included 1.4 to 3.3 times more species than the most species-rich community of that gradient, depending on the plant or animal taxon considered (Novotny & Toko, 2015).
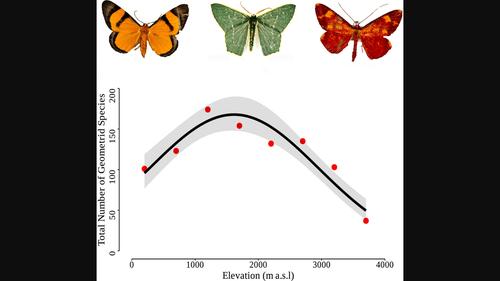
Insects, with their high species richness and diverse ecological functions, have often been used as model taxa to study ecological trends along elevational gradients (Chen et al., 2009; Colwell et al., 2008; McCain, 2009, 2010). Geometrid moths (Geometridae, Lepidoptera) have become a widely used model for insect community studies (Beck et al., 2017) because they are one of the largest, but still taxonomically manageable, insect families, with ~24,000 described species (Rajaei et al., 2022). In addition, adults can be easily surveyed with light traps. Because of their herbivorous lifestyle, geometrids are sensitive to environmental changes, including climate change (Chen et al., 2009), vegetation disturbance (Holloway et al., 1992) and successional dynamics of vegetation (Ashton et al., 2016).
Geometrids generally exhibit maximum diversity at mid-elevations along elevational gradients. However, despite the wealth of available data, the causes of this pattern are not clear (Beck et al., 2017). Here, we study highly diverse geometrid communities along a complete rainforest elevational gradient at Mt. Wilhelm (Papua New Guinea) with the goal of describing and explaining elevational trends in their species composition and alpha and beta diversity and characterising the contribution of elevational gradients to regional gamma diversity. We hypothesise that a unimodal maximum in geometrid species diversity is a composite result of decreasing abiotic environment favourability, particularly decreasing temperature, and increasing biotic favourability, particularly decreasing predation pressure, with increasing elevation. In addition, we also expect a positive correlation of species diversity of mostly herbivorous geometrid moths with the diversity of vegetation that constitutes their resource base. These factors, together with plant species composition, should also explain the species composition of geometrid communities. Finally, we expect increasing beta diversity with increasing elevation as a consequence of Rapoport's rule.
中文翻译:
巴布亚新几内亚完整雨林海拔梯度上蛾群落(尺蛾科、鳞翅目)的生态趋势
介绍
全球生物多样性最大值,例如,包括世界上所有六个植物区系最丰富的地区,每 10,000 km 2拥有超过 5000 种植物物种(Kier 等人, 2005 年)),通常包括潮湿热带地区的长海拔梯度,特别是从低地延伸到林线的“完整”雨林海拔梯度,通常为~3700 m asl。这些梯度在一个小地理区域内涵盖了一系列高度多样化的雨林生态系统,这是由于沿陡峭的非生物和生物生态梯度的高物种更替造成的。这些样线是研究群落组装机制的理想模型,而不会使扩散限制的影响复杂化。最近,它们在气候变化研究中也变得很重要,因为它们在有限的区域内再现了广泛的温度范围。
初步估计,物种多样性要么随着海拔的增加而单调减少,要么在中间海拔处达到最大值(Rahbek, 1995;Stevens, 1992)。物种多样性的单调下降可能直接或间接归因于温度和/或土地面积随海拔升高而降低(Beck&Chey, 2008;Beck&Kitching, 2009)。其他重要因素包括初级生产力和栖息地复杂性,包括对鸟类重要的植被结构(Sam 等, 2019)或对蚂蚁重要的垃圾数量(Moses 等, 2021;Pérez-Toledo 等, 2021 ))。中海拔的最大多样性很少对应于单个非生物或生物驱动变量。它可能是中域效应(Colwell et al., 2016)和/或多种因素组合的产物。特别是,随着海拔的升高,来自捕食者和病原体的生物压力往往会降低,而非生物胁迫往往会增加(Péré等, 2013)。不幸的是,人们对食草、寄生或捕食等营养相互作用强度的趋势知之甚少,因为它们通常是由很少一起研究的多个类群引起的(Bärtschi 等人,2019 年;Peters 等人, 2016年 )。
整个海拔梯度的伽马多样性是各个群落的阿尔法多样性值和跨海拔物种周转率的乘积。拉波波特规则表明,β 多样性应随着海拔的增加而减少,因为山地物种应比低地物种具有更大的海拔范围(Stevens, 1992)。然而,对沿海拔梯度的 β 多样性趋势的研究少于 α 多样性趋势(Grytnes & McCain, 2007)。沿海拔梯度的β多样性决定了区域、横断面伽玛多样性的总体范围。例如,新几内亚威廉山从低地雨林到林线的完整雨林海拔梯度包含的物种比该梯度中物种最丰富的群落多 1.4 至 3.3 倍,具体取决于所考虑的植物或动物分类群(Novotny & Toko) , 2015)。
昆虫因其物种丰富度高、生态功能多样,常被用作模型类群来研究沿海拔梯度的生态趋势(Chen et al., 2009; Colwell et al., 2008 ; McCain , 2009 , 2010 )。尺蛾(尺蛾科、鳞翅目)已成为昆虫群落研究广泛使用的模型(Beck 等人, 2017),因为它们是最大的昆虫科之一,但在分类学上仍然易于管理,有约 24,000 个描述的物种(Rajaei 等人) ., 2022 )。此外,可以使用光陷阱轻松地调查成年人。由于尺蠖的食草生活方式,它们对环境变化很敏感,包括气候变化(Chen et al., 2009)、植被干扰(Holloway等, 1992)和植被演替动态(Ashton等, 2016)。
尺蠖通常在沿海拔梯度的中海拔处表现出最大的多样性。然而,尽管有大量可用数据,但这种模式的原因尚不清楚(Beck 等人, 2017)。在这里,我们沿着威廉山(巴布亚新几内亚)的完整雨林海拔梯度研究高度多样化的尺蛾群落,目的是描述和解释其物种组成以及α和β多样性的海拔趋势,并表征海拔梯度对区域的贡献伽马多样性。我们假设,尺蛾物种多样性的单峰最大值是随着海拔升高,非生物环境有利性降低(特别是温度降低)和生物有利性增加(特别是捕食压力降低)的复合结果。此外,我们还预计大多数草食性尺蛾的物种多样性与构成其资源基础的植被多样性呈正相关。这些因素,与植物物种组成一起,还应解释尺蠖群落的物种组成。最后,我们预计,由于拉波波特规则,β 多样性会随着海拔的增加而增加。





































 京公网安备 11010802027423号
京公网安备 11010802027423号