Archives of Pharmacal Research ( IF 6.9 ) Pub Date : 2023-01-06 , DOI: 10.1007/s12272-023-01426-5
Ga-Eun Lee 1 , Dohyun Jeung 1 , Weidong Chen 1 , Jiin Byun 1 , Joo Young Lee 1 , Han Chang Kang 1 , Hye Suk Lee 1 , Dae Joon Kim 2 , Jin-Sung Choi 3 , Cheol-Jung Lee 1, 4 , Hyun-Jung An 1 , Yong-Yeon Cho 1, 5
|
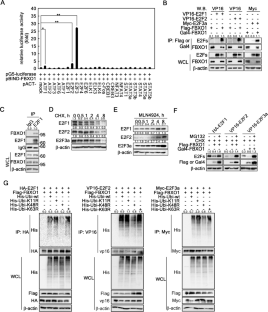
E2F 1、2 和 3a(简称 E2F)是 E2F 转录因子家族的一个亚家族,在细胞周期进程、DNA 复制、DNA 修复、细胞凋亡和分化中发挥重要作用。尽管 E2Fs 的转录调控主要集中在口袋蛋白视网膜母细胞瘤蛋白复合物上,但最近的研究表明 E2Fs 的翻译后修饰和稳定性调控在多种细胞过程中起着关键作用。在这项研究中,我们发现 FBXO1 是 S 期激酶相关蛋白 1 (SKP1)-cullin 1-F-box 蛋白 (SCF) 复合物的一个组成部分,是 E2Fs 结合伙伴。此外,FBXO1 与 E2Fs 的结合诱导了 K48 泛素化和随后的 E2Fs 蛋白酶体降解。结合域分析表明,位于 E2F 的二聚化域中的 Arg (R)/Ile (I) 和 R/Val (V) 基序,E2F 1 和 3a 以及 E2F2 分别充当 FBXO1 的降解决定子基序 (DM)。值得注意的是,DM 中的 RI/AA 或 RV/AA 突变减少了 FBXO1 介导的泛素化并延长了 E2F 的半衰期。重要的是,E2F 的稳定性受到位于 DM 的 RI 和 RV 残基附近的苏氨酸残基磷酸化的影响。磷酸化预测数据库分析和特异性抑制剂分析表明,MEK/ERK 信号分子在 FBXO1/E2Fs 的相互作用和调节 E2F 蛋白转换中起着关键作用。此外,通过敲低 FBXO1 提高了 E2Fs 蛋白水平,通过 sh-E2F3a 延迟 G 降低了 E2Fs 蛋白水平 E2F 的稳定性受位于 DM 的 RI 和 RV 残基附近的苏氨酸残基磷酸化的影响。磷酸化预测数据库分析和特异性抑制剂分析表明,MEK/ERK 信号分子在 FBXO1/E2Fs 的相互作用和调节 E2F 蛋白转换中起着关键作用。此外,通过敲低 FBXO1 提高了 E2Fs 蛋白水平,通过 sh-E2F3a 延迟 G 降低了 E2Fs 蛋白水平 E2F 的稳定性受位于 DM 的 RI 和 RV 残基附近的苏氨酸残基磷酸化的影响。磷酸化预测数据库分析和特异性抑制剂分析表明,MEK/ERK 信号分子在 FBXO1/E2Fs 的相互作用和调节 E2F 蛋白转换中起着关键作用。此外,通过敲低 FBXO1 提高了 E2Fs 蛋白水平,通过 sh-E2F3a 延迟 G 降低了 E2Fs 蛋白水平1 /S 细胞周期转换,导致癌细胞增殖受到抑制。这些结果表明,FBXO1-E2Fs 轴介导的精确 E2Fs 稳定性调节通过 G 1 /S 细胞周期转换在细胞增殖中起关键作用。
"点击查看英文标题和摘要"
































 京公网安备 11010802027423号
京公网安备 11010802027423号