Our official English website, www.x-mol.net, welcomes your
feedback! (Note: you will need to create a separate account there.)
金黄色葡萄球菌广谱 β-内酰胺耐药的结构基础
Nature ( IF 50.5 ) Pub Date : 2023-01-04 , DOI: 10.1038/s41586-022-05583-3 J Andrew N Alexander 1, 2 , Liam J Worrall 1, 2, 3 , Jinhong Hu 1, 2 , Marija Vuckovic 1, 2 , Nidhi Satishkumar 4, 5 , Raymond Poon 4, 5 , Solmaz Sobhanifar 1, 2 , Federico I Rosell 1, 2 , Joshua Jenkins 1, 2 , Daniel Chiang 6 , Wesley A Mosimann 1, 2 , Henry F Chambers 7 , Mark Paetzel 6 , Som S Chatterjee 4, 5 , Natalie C J Strynadka 1, 2, 3
Affiliation
|
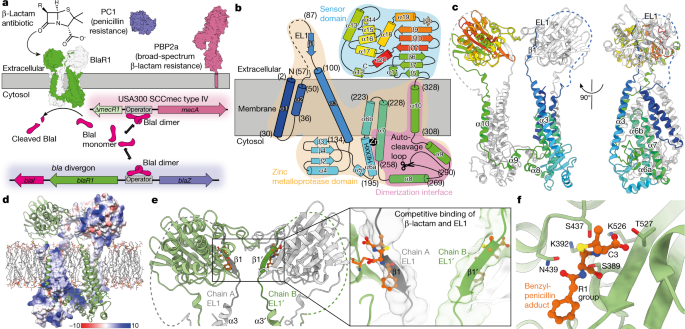
金黄色葡萄球菌的广谱 β-内酰胺类抗生素耐药性是一项全球医疗保健负担1,2。在临床菌株中,耐药性主要受 BlaR1 3 控制,BlaR13 是一种受体,通过其传感器结构域的酰化感知 β-内酰胺,诱导跨膜信号传导和面向细胞质的金属蛋白酶结构域4 的激活。金属蛋白酶结构域在 BlaI 去抑制中发挥作用,诱导 blaZ(β-内酰胺酶 PC1)和 mecA(β-内酰胺抗性细胞壁转肽酶 PBP2a)表达3,4,5,6,7。在这里,克服了孤立的障碍,我们表明 BlaR1 直接切割 BlaI,这是灭活的需要,不需要前面建议的附加组分8。BlaR1(野生型和自切割缺陷型 F284A 突变体,有或没有 β-内酰胺)的冷冻电子显微镜结构揭示了一个结构域交换的二聚体,我们认为这对内部信号环的稳定至关重要。BlaR1 在 Ser283 和 Phe284 之间的顺式中经历自发自剪,我们描述了自身和 BlaI 切割的催化机制和特异性。这些结构表明,变构信号源于 β-内酰胺诱导的排除在传感器域活性位点竞争性结合的突出细胞外环,驱动随后的动态运动,包括传感器向膜的移动以及伴随锌金属蛋白酶结构域的变化。我们建议这增强了自裂解产物从活性位点的排出,将平衡转变为允许有效 BlaI 切割的状态。 总的来说,本研究提供了一种双组分信号受体的结构,该受体通过抑制物的直接切割介导作用——在这种情况下,抗生素耐药性。
"点击查看英文标题和摘要"






























 京公网安备 11010802027423号
京公网安备 11010802027423号